5018
The detrimental lifelong impacts of early life adversity on brain1Laboratory of Brain Connectomics, RIKEN BDR, Kobe, Japan, 2Nara Woman’s University, Kobe, Japan
Synopsis
Keywords: Psychiatric Disorders, Brain
Motivation: Early life adversities (ELAs) have long-lasting detrimental impacts on physical and mental health across life course, insisting the experience of ELAs disturb healthy neuronal brain development.
Goal(s): Our goal is to investigate on the underlying mechanism of the long-lasting ELAs impacts on brain.
Approach: We examined the brains of non-human primate experienced with the ELAs of social separation from conspecific caregivers from our non-human primate MRI database in retrospective manner.
Results: Non-human primate brain with caregiver adversities (i.e. ELAs) exerts underdevelopment of multiple brain regions in both cortical and subcortical regions.
Impact: Our study demonstrates the lifelong detrimental impact of the early life adversities on brains, proving that the early life experiences, especially social ones, are the important factor for structuring brain during development.
Introduction
Early life adversities (ELAs) are the exposures to negative experiences at early life stage. There has been scientific dogma that ELAs has long-lasting effects on both physical and mental health across life course. Common marmoset (Callithrix jacchus) is a highly social primate, and all the family members actively participates caregiving in offspring. However, sometimes, marmosets exhibit neglect and abusive behaviors toward their offspring. In such cases, our lab applied artificial rearing (AR). Consequently, baby marmosets experience separation from their family and lost expected conspecific parental care. In this study, we defined AR marmoset as a model for ELAs and examined the structural brain differences between AR and parental rearing (PR) adult marmosets in a retrospective manner.Methods
From the Japan’s national projects (Brain/MINDS and Brain/MINDS-beyond) 1 launched, we have been collected marmoset brain images with a 3.0-Tesla MRI scanner (3-Tesla, MAGNETOME Prisma, Siemens AG, Erlangen, Germany) with a 16ch marmoset head coil (Takashima Seisakusho Co., Ltd., Tokyo, Japan) with actively shielded gradients at a maximum strength of 80 mT/m. The structural brain dataset, including T1-weighted (T1w) and T2-weighted (T2w) images, in the same scanning protocol contained 97 marmosets (66 males; 6.2±2.6 years old) , and sixteen marmosets (15 males) out of them were artificially reared by the experimenters (C.T. and A.K.) due to their parents’ abuse (filial cannibalism), neglect, or poor physical condition after delivery. For data processing, concerning recent practice of data sharing in human research field, our current dataset is organized in the Brain Imaging Data Structure (BIDS)2 format and processed by human connectome project (HCP) pipeline3 customized for non-human primates4. Then, statistical regression analysis of brain volumes, thickness, and area derived from FreeSurfer version 7.4.0 were performed to see the effect of AR (see Figure.1) using a statistical software R 4.03.Results
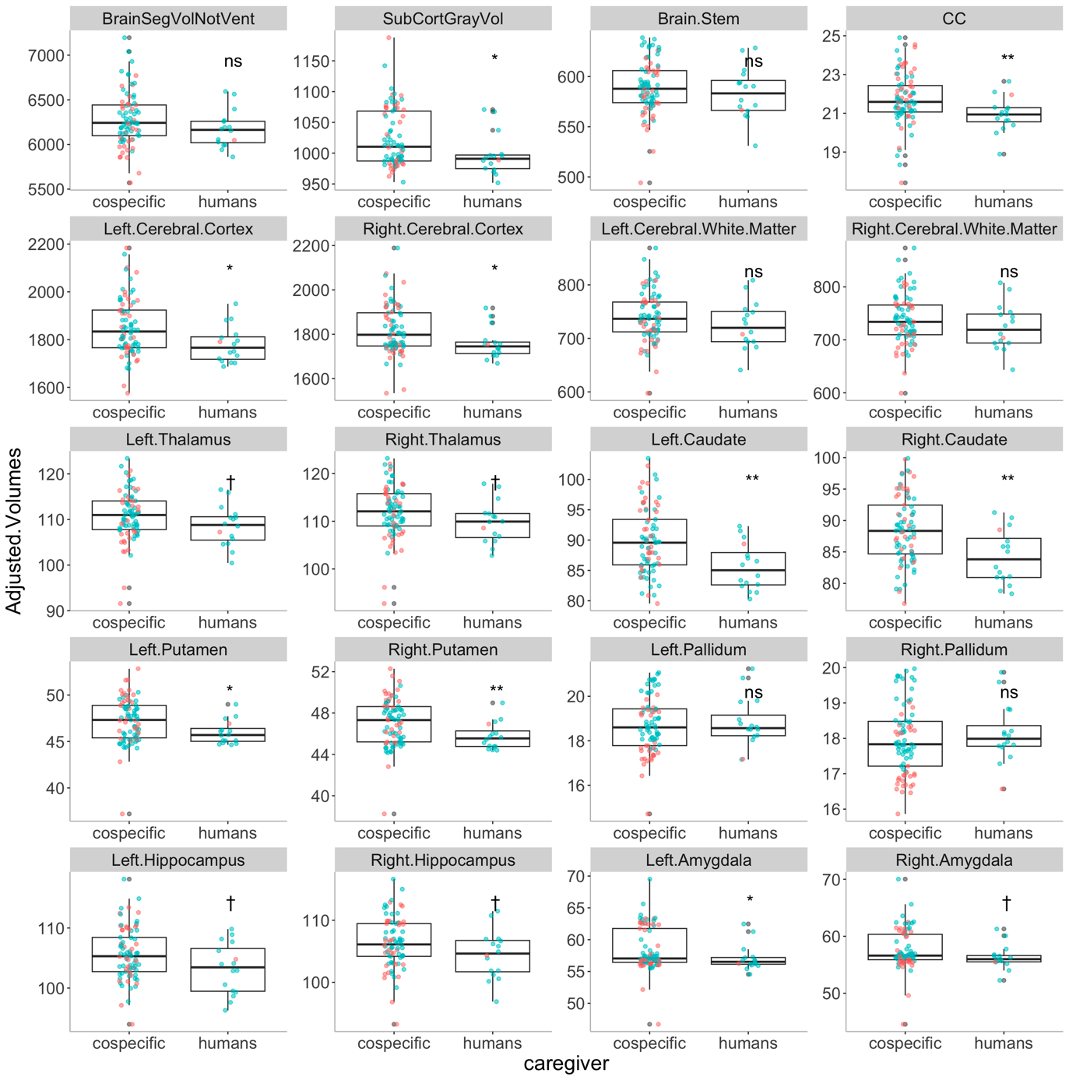
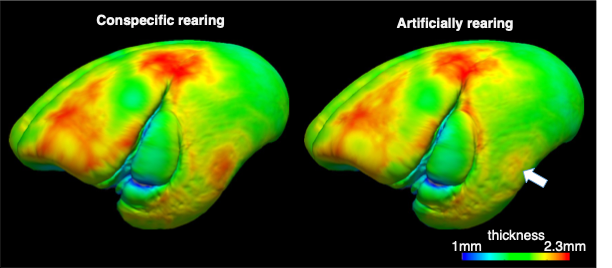
Because this study is retrospective, there was significant age difference between PR and AR groups (t(96)=-2.20, p = 0.04 ). In addition, estimated intracranial volumes were also significantly smaller in AR group (t(96)=-2.58,p= 0.017) despite no significant body size difference ((t(96)=-0.1, p = 0.92 ). After regressing out these effects, we found that multiple brain regions were significantly smaller in AR compared to PR (Figure. 2), insisting the underdevelopment of those brain regions. The most robust differences between AR and PR were observed in claustrum (i.e. bilateral caudate and putamen) and corpus callosum. In addition to the smaller volumes of claustrum, thinner thickness of middle temporal regions (see the arrow in Figure 3), those of which regions play important roles of motor functions.Discussions
In this study, we examined the brain of artificially reared adult marmosets as a model experienced the early life caregiver adversities. Our results demonstrated underdevelopment of multiple supratentorial regions, especially motor-function related areas, which suggest that receiving external stimuli via parent-child interactions and/or belong to the social community plays important role on brain structures in later life stage. Indeed, psychological well-beingness in adulthood has been said to be partially based childhood experiences in both humans and non-human primates. Moreover, artificially reared common marmosets showed changes in social behavior and brain serotonin functions5. Early life stage is the sensitive period for brain forming and refining complex neural circuits through synaptogenesis, synaptic pruning, myelination, and neurogenesis. According to the review6, Brain in this period is also sensitive to the ELAs-induced stress. Stress disrupts proper neuronal development, causing long-lasting and pervasive effects on neural circuit maturation.Interestingly, the regions that showed either significant or tendency for underdevelopment, such as corpus callosum, hippocampus, and amygdala, are the regions consistently reported in the ELAs studies, especially the adversity of social separation from caregivers, regardless of species including human beings. Thus, the impact of the ELAs on those brain regions would be conserved across social animals and could be potential target for early intervention or treatment of psychopathologies originated from the ELAs. As future directions, because in our database some marmosets also have serotonin receptor PET image and personality test, we would examine the ELA effects from multiple perspectives to unveil its underlying mechanisms.Acknowledgements
This research was supported by Agency for Medical Research and development (AMED) [grant number: JP18dm0307006].References
- Nature Publishing Group. (n.d.). Focal Point on Brain Science in Japan. Nature news. https://www.nature.com/collections/bbcgefaifg
- Gorgolewski, K. J., Auer, T., Calhoun, V. D., Craddock, R. C., Das, S., Duff, E. P., ... & Poldrack, R. A. (2016). The brain imaging data structure, a format for organizing and describing outputs of neuroimaging experiments. Scientific data, 3(1), 1-9.
- Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S., Fischl, B., Andersson, J. L., ... & Wu-Minn HCP Consortium. (2013). The minimal preprocessing pipelines for the Human Connectome Project. Neuroimage, 80, 105-124.
- Hayashi, T., Hou, Y., Glasser, M. F., Autio, J. A., Knoblauch, K., Inoue-Murayama, M., ... & Van Essen, D. C. (2021). The nonhuman primate neuroimaging and neuroanatomy project. Neuroimage, 229, 117726.
- Yokoyama, C. et al., “Influence of early social experience on sociability and brain serotonin function in common marmosets” (2015) Society for Neuroscience Annual Meeting, 2015 Abstract
- Malave, L., van Dijk, M. T., & Anacker, C. (2022). Early life adversity shapes neural circuit function during sensitive postnatal developmental periods.
Figures
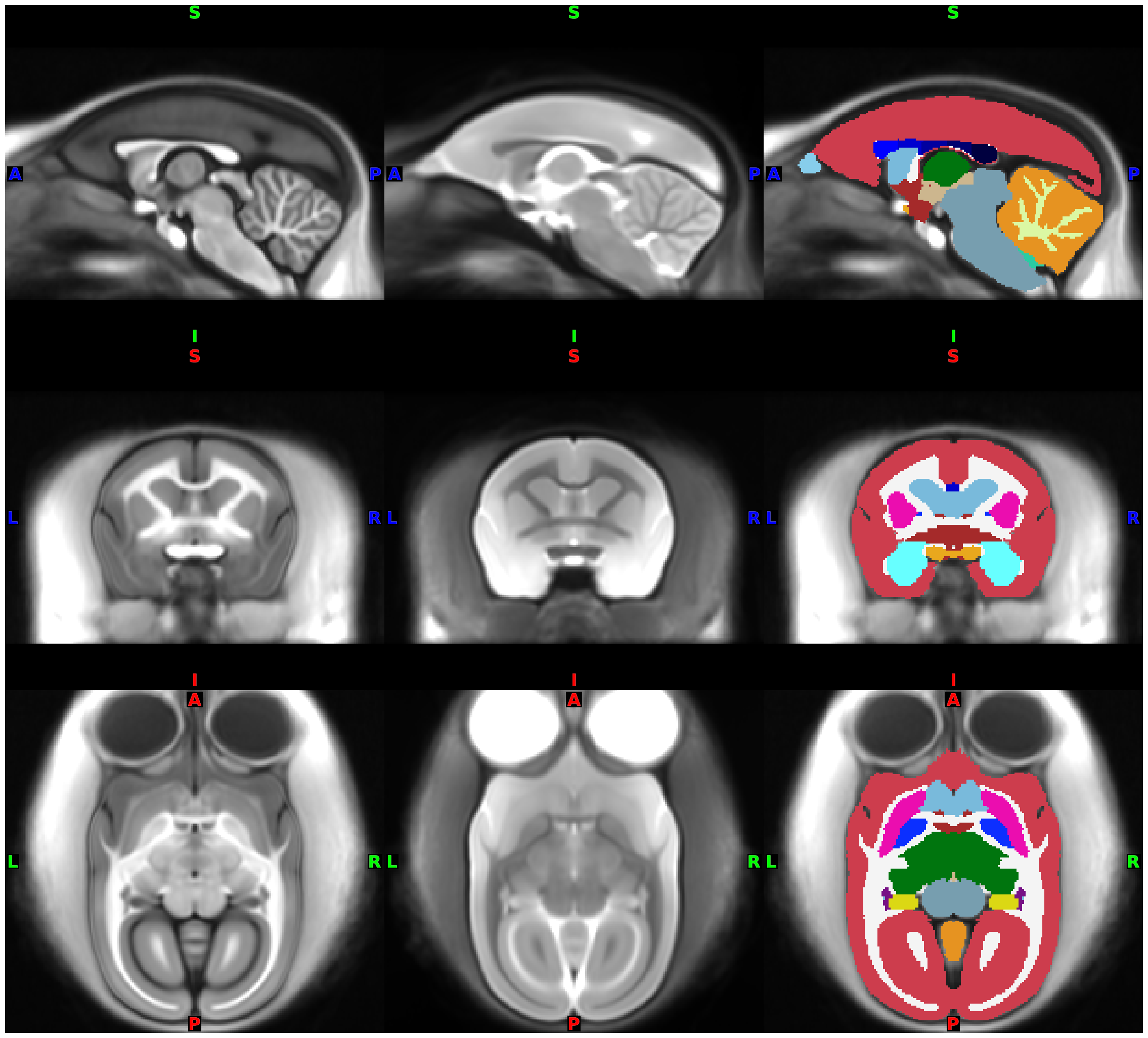
Derived data from FreeSurfer:
0.2mm isotropic (set –hires option at recon-all) averaged T1.mgz, T2.norm.mgz, and aseg.presurf.mgz