4975
Time course of structural neuroplasticity induced by procedural motor learning: DTI vs SANDI1IFIBIO-Houssay, Department of Physiology, School of Medicine, University of Buenos Aires, Argentina, Buenos Aires, Argentina, 2Brain Research Imaging Centre (CUBRIC), School of Psychology and School of Computer Science and Informatics, Cardiff University, Cardiff, UK., Cardiff, United Kingdom, 3Athinoula A. Martinos Center for Biomedical Imaging, Department of Radiology, Massachusetts General Hospital, Harvard Medical School, Boston, MA, USA., Boston, MA, United States
Synopsis
Keywords: Gray Matter, Neuroscience, motor learning, plasticity, SANDI
Motivation: Performance gains during motor sequence learning (MSL) are linked to increased hippocampal activity and fast changes in mean diffusivity. Yet, DTI is not biologically specific.
Goal(s): To identify the biological compartment(s) driving changes in gray-matter microstructure during MSL.
Approach: We applied SANDI to multi-shell-DWI acquired in 28 subjects on the Connectome scanner before, 30 min, and 24h post MSL training.
Results: MSL-induced transient changes in DTI were associated with a temporary increase in soma fraction at 30 min, compatible with a fast homeostatic response (e.g. astrocytic swelling). Long-lasting DTI changes were associated with an increment in neurite fraction compatible with structural remodelling.
Impact: Our work may broaden the scope of understanding of human hippocampal memory and help pinpoint the biological substrates of plasticity. Moreover, it may set the basis for developing a biologically meaningful biomarker of neuroplasticity to detect early signs of neuropathology.
Introduction
The most important computation attributed to the human hippocampus is episodic memory. Nevertheless, recent studies have linked this structure to non-declarative memory (1-3). We have shown that improvements in performance during motor sequence learning (MSL) are associated with an increment in hippocampal and precuneus activity (4), and rapid alterations in microstructure in the same regions quantified using mean diffusivity (MD). Reductions in MD are hypothesized to reflect a decrease in interstitial space accompanying astroglial expansion during synaptic plasticity (5). Yet, diffusion tensor imaging (DTI) reflects a combination of intra- and extracellular water diffusion effects that limits its specificity to detect changes at the cellular level and its potential as a biologically meaningful proxy for structural plasticity. To overcome this limitation, here, we used the strong gradients of the original Connectome MRI scanner with a compartment-based model for apparent cell soma and neurite density imaging (SANDI)(6) to identify the biological compartment(s) driving changes in gray-matter microstructure during MSL.Methods
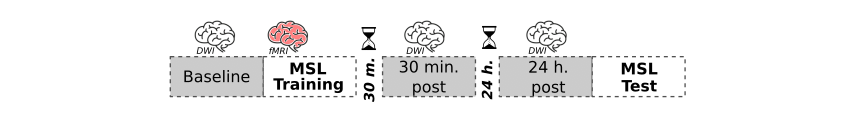
Twenty-eight healthy human subjects (aged 18-35 y/o; 15 female) learned an MSL task, which required executing a 5-item sequence of finger movements on a keyboard using four fingers of the non-dominant hand. Functional, T1-weighted, and Diffusion-weighted MRI (DWI) images were acquired on the Connectome MRI scanner (MAGNETOM Connectom, Siemens Healthineers, Erlangen, Germany). Functional images were obtained following a block design during MSL (15 blocks of self-paced task alternated with 25s rest), and were pre-processed and analysed as in our previous study (4). Multi-shell DWI were acquired following a longitudinal design before learning (baseline), 30 min and 24 h post learning (Fig 1) to track changes in microstructure compatible with short- and long- term plasticity: SMS=2, voxel size: 2×2×2 mm, 32 directions of b-values=200, 800, 1200, 2400 s/mm2, and 64 directions of b-values=4000, 6000, 8000 s/mm2, 22 b0 volumes, FOV=216 mm. DWI images were first denoised, corrected for nonlinearity of gradients, geometric distortions, head motion, eddy currents, and b-vector, and then modelled using DTI and SANDI. For each subject and each voxel, MD maps were computed using the 1200 s/mm2 shell; on the other hand SANDI was fit on the multi-shell DWI acquisition, obtaining diffusion fractions corresponding to the extracellular (FEXTRA), intraneurite (FNEURITE), and soma (FSOMA) compartments. MD maps and SANDI's fractions were then transformed to stereotaxic space (MNI152) using an ANTs-based pipeline (7). Whole-brain longitudinal MD changes across sessions were statistically assessed using a non-parametric clusterwise approach (cluster forming threshold=0.001) implemented in the Sandwich Estimator (SwE) toolbox (p<0.05 FWE-corrected)(8). The clusters from this analysis were used as ROIs to assess differences across sessions for the FEXTRA, FNEURITE and FSOMA fractions (ANOVA, p<0.05 corrected by Bonferroni). In addition, both DTI and SANDI were computed independently in the posterior, body and anterior hippocampi segmented based on each subject's T1 (9).Results
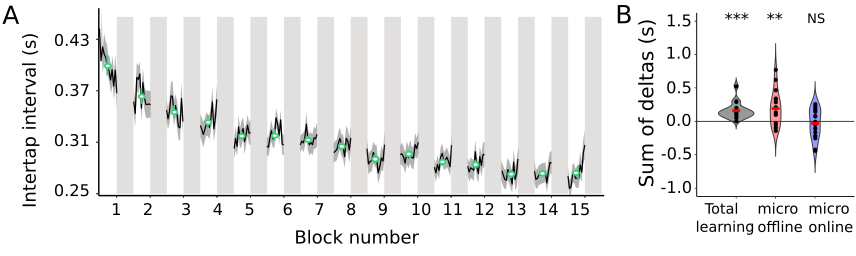
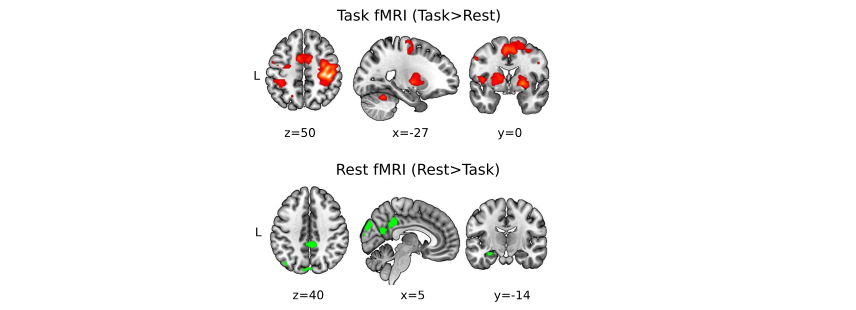
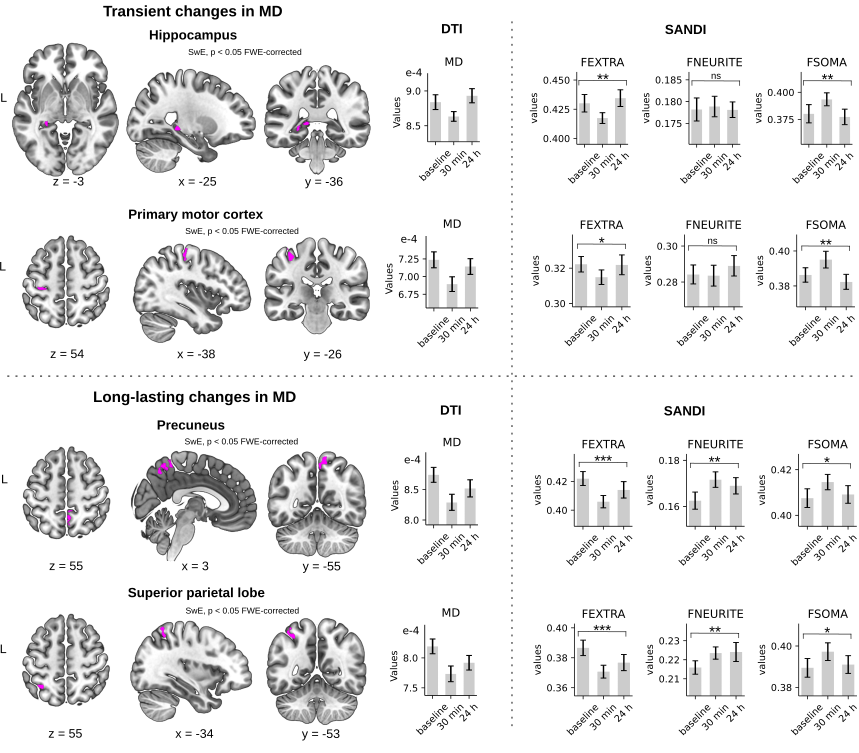
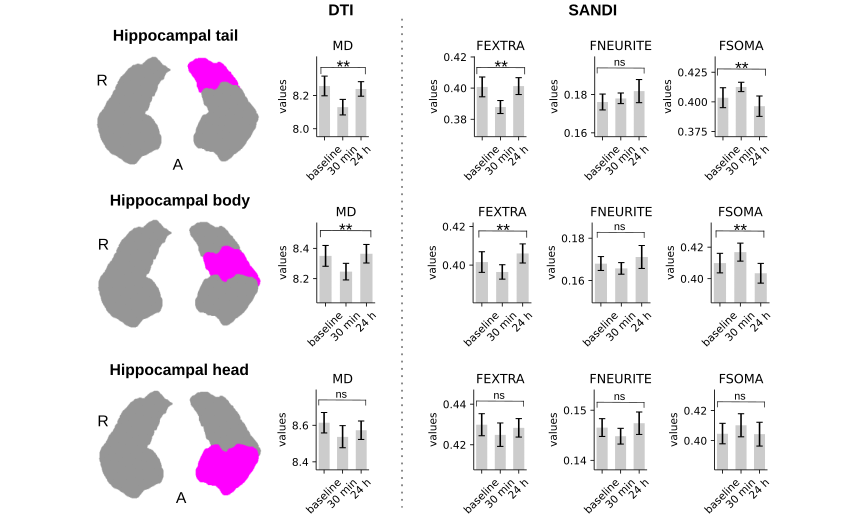
In line with our previous study (4), we found: i) that improvements in performance on the MSL task took place during the rest periods interleaved with practice, known as micro-offline gains (MOGs, Fig 2), ii) that activity in the left hippocampus and the right precuneus increased during the rests periods interleaved with practice (Fig 3), and iii) that MSL decreased MD in the left posterior hippocampus and right precuneus (Fig 4, top and bottom left panels). Moreover, we detected further a decrease in MD in the left primary motor (M1) and posterior parietal cortex (PPC), two regions that participate in this type of motor learning. While MD changes were transient in the hippocampus and M1, returning to baseline values at 24 h, MD in the precuneus and PPC were long-lasting (Fig 4; left panel). Importantly, transient decreases in MD were associated with a temporary increase in FSOMA at 30 min, whereas long-lasting MD changes were associated with an increment in FNEURITE (Fig 4, right panels). Critically, comparison of DTI and SANDI metrics obtained independently from each hippocampal segment yielded anatomically consistent changes in microstructure across the body and tail of the left hippocampus (Fig 5). No changes were detected for either metric in the head or in the right hippocampus.Conclusion and Discussion
We conclude that MSL induces a non-specific -transient- increase in the soma compartment of the hippocampus and other cortical regions involved in MSL, and a long-lasting increase in the neurite fraction of the precuneus and the PPC. Whereas the former may reflect a fast homeostatic response such as astrocytic swelling associated with LTP-like processes (5, 10-11), the latter may be compatible with structural remodelling of astrocytes and/or neurons.Acknowledgements
We thank the Argentinian Agency for the Promotion of Science and Technology (FONCyT: PICT 2018-1150; PICT 2019-2156) for their financial support.References
1. Döhring, J., Stoldt, A., Witt, K., Schönfeld, R., Deuschl, G., Born, J., & Bartsch, T. (2017). Motor skill learning and offline-changes in TGA patients with acute hippocampal CA1 lesions. Cortex, 89, 156-168.
2. Long, J., Feng, Y., Liao, H., Zhou, Q., & Urbin, M. A. (2018). Motor sequence learning is associated with hippocampal subfield volume in humans with medial temporal lobe epilepsy. Frontiers in Human Neuroscience, 12, 367.
3. Schapiro, A. C., Reid, A. G., Morgan, A., Manoach, D. S., Verfaellie, M., & Stickgold, R. (2019). The hippocampus is necessary for the consolidation of a task that does not require the hippocampus for initial learning. Hippocampus, 29(11), 1091-1100.
4. Jacobacci, F., Armony, J. L., Yeffal, A., Lerner, G., Amaro Jr, E., Jovicich, J., ... & Della-Maggiore, V. (2020). Rapid hippocampal plasticity supports motor sequence learning. Proceedings of the National Academy of Sciences, 117(38), 23898-23903.
5. Sagi, Y., Tavor, I., Hofstetter, S., Tzur-Moryosef, S., Blumenfeld-Katzir, T., & Assaf, Y. (2012). Learning in the fast lane: new insights into neuroplasticity. Neuron, 73(6), 1195-1203.
6. Palombo, M., Ianus, A., Guerreri, M., Nunes, D., Alexander, D. C., Shemesh, N., & Zhang, H. (2020). SANDI: a compartment-based model for non-invasive apparent soma and neurite imaging by diffusion MRI. Neuroimage, 215, 116835.
7. Jacobacci, F., Jovicich, J., Lerner, G., Amaro Jr, E., Armony, J. L., Doyon, J., & Della‐Maggiore, V. (2020). Improving spatial normalization of brain diffusion MRI to measure longitudinal changes of tissue microstructure in the cortex and white matter. Journal of Magnetic Resonance Imaging, 52(3), 766-775.
8. Guillaume, B., Hua, X., Thompson, P. M., Waldorp, L., Nichols, T. E., & Alzheimer's Disease Neuroimaging Initiative. (2014). Fast and accurate modelling of longitudinal and repeated measures neuroimaging data. Neuroimage, 94, 287-302.
9. Iglesias, J. E., Augustinack, J. C., Nguyen, K., Player, C. M., Player, A., Wright, M., ... & Alzheimer's Disease Neuroimaging Initiative. (2015). A computational atlas of the hippocampal formation using ex vivo, ultra-high resolution MRI: application to adaptive segmentation of in vivo MRI. Neuroimage, 115, 117-137.
10. Jin BJ, Zhang H, Binder DK, Verkman AS. Aquaporin-4-dependent K(+) and water transport modeled in brain extracellular space following neuroexcitation. J Gen Physiol. 2013;141(1):119-32. Doi: 10.1085/jgp.201210883.
11. Mader S, Brimberg L. Aquaporin-4 Water Channel in the Brain and Its Implication for Health and Disease. Cells. 2019;8(2). Epub 20190127. doi: 10.3390/cells8020090.
Figures