4719
Particle-based modelling of isotope exchange at equilibrium in hyperpolarized [1-13C]pyruvate MR1Medical Biophysics, University of Toronto, Toronto, ON, Canada, 2Physical Sciences, Sunnybrook Research Institute, Toronto, ON, Canada
Synopsis
Keywords: Biology, Models, Methods, Modelling
Motivation: Isotope exchange at equilibrium complicates interpretation hyperpolarized [1-13C]pyruvate MR. Modelling this phenomenon could help to quantify the effect of exchange on HP MR signal.
Goal(s): Investigate how isotope exchange (compensatory backward conversion of unlabeled lactate concurrent with labeled lactate production) occurs and affects hyperpolarized [1-13C]pyruvate MR.
Approach: Develop a realistic computational model of pyruvate-lactate interconversion and apply it to characterize how different levels of endogenous lactate influence metabolic reaction kinetics and exchange.
Results: Elevation of unlabeled lactate increases isotope exchange at equilibrium. Net production of lactate occurs unless total lactate exceeds its equilibrium ratio with total pyruvate.
Impact: This in silico model of isotope exchange in hyperpolarized [1-13C]pyruvate MR realistically replicates spectroscopic measurements, with particle-level data for a range of conditions providing insight into metabolic dynamics relevant to complex cellular architectures with different local equilibria.
Introduction
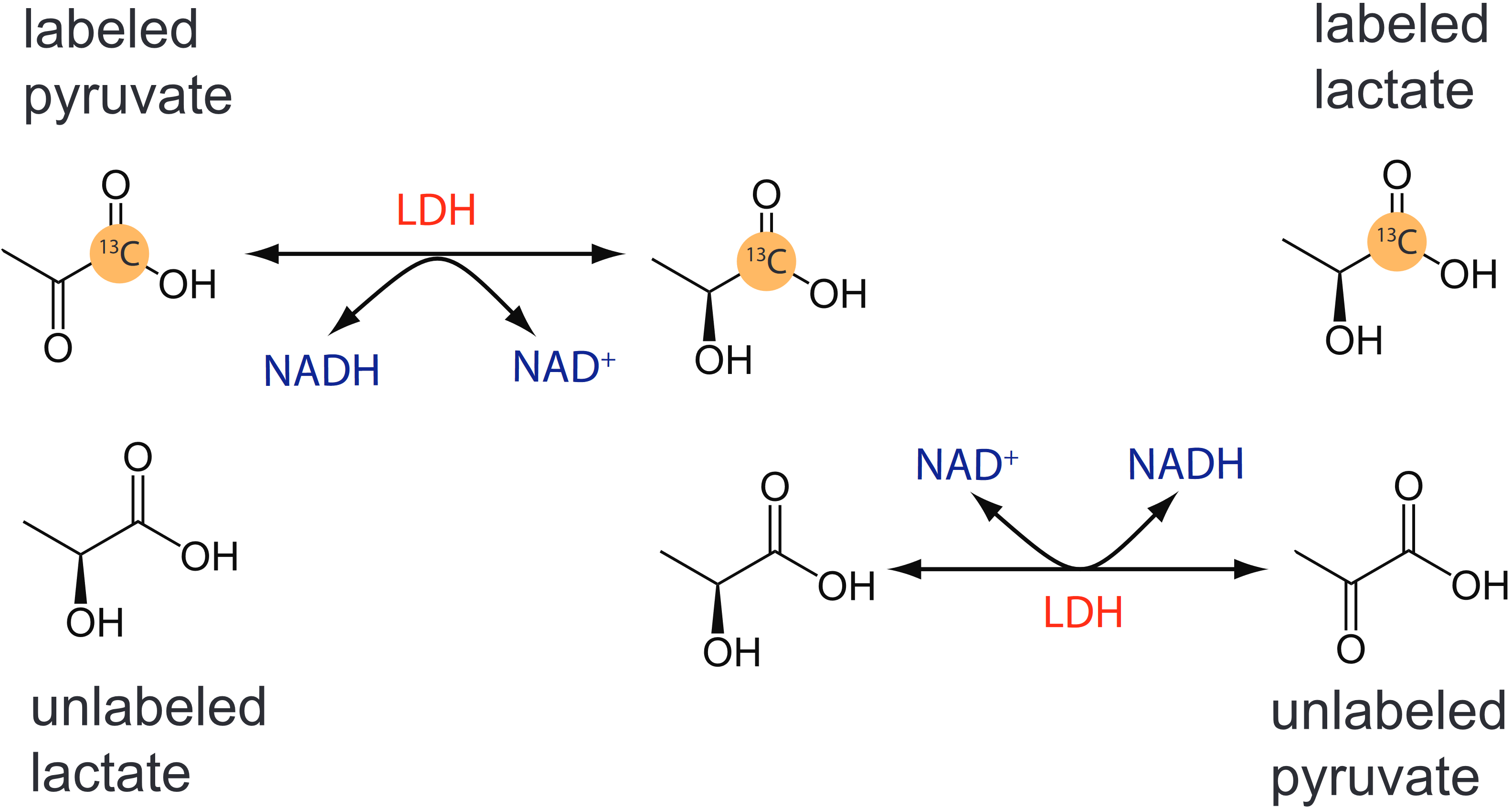
It is well known that the pyruvate and lactate within the cytosol can exchange between one another through the two-way reaction catalyzed by the LDH enzyme family. This can confound interpretation of 13C-metabolite signals due to the phenomenon of “isotope exchange at equilibrium”.1 In this scenario, [1-13C]lactate production occurs alongside compensatory reactions involving the unlabeled metabolite pools, resulting in zero net flux from pyruvate to lactate, as depicted in Figure 1. Homeostatic equilibrium generally favors a high ratio of lactate to pyruvate,2–4, but hyperpolarized 13C MR (HP 13C MR) perturbs this state through injection of 250 mM [1-13C]pyruvate, supplying potentially supraphysiological levels of exogenous pyruvate to cells.5,6 Since HP 13C MR signals only reflect labeled metabolites, the degree to which metabolic flux occurs is unclear; forward conversion reactions of [1-13C]pyruvate may be counterbalanced by exchange with the endogenous metabolite pool. In this study, we tested whether a particle-based in silico model of pyruvate-lactate interconversion would reproduce isotope exchange at equilibrium, making it useful for understanding the effect of this exchange phenomenon in more complex cell architectures involving cellular compartments with different equilibria, such as the human brain.Methods
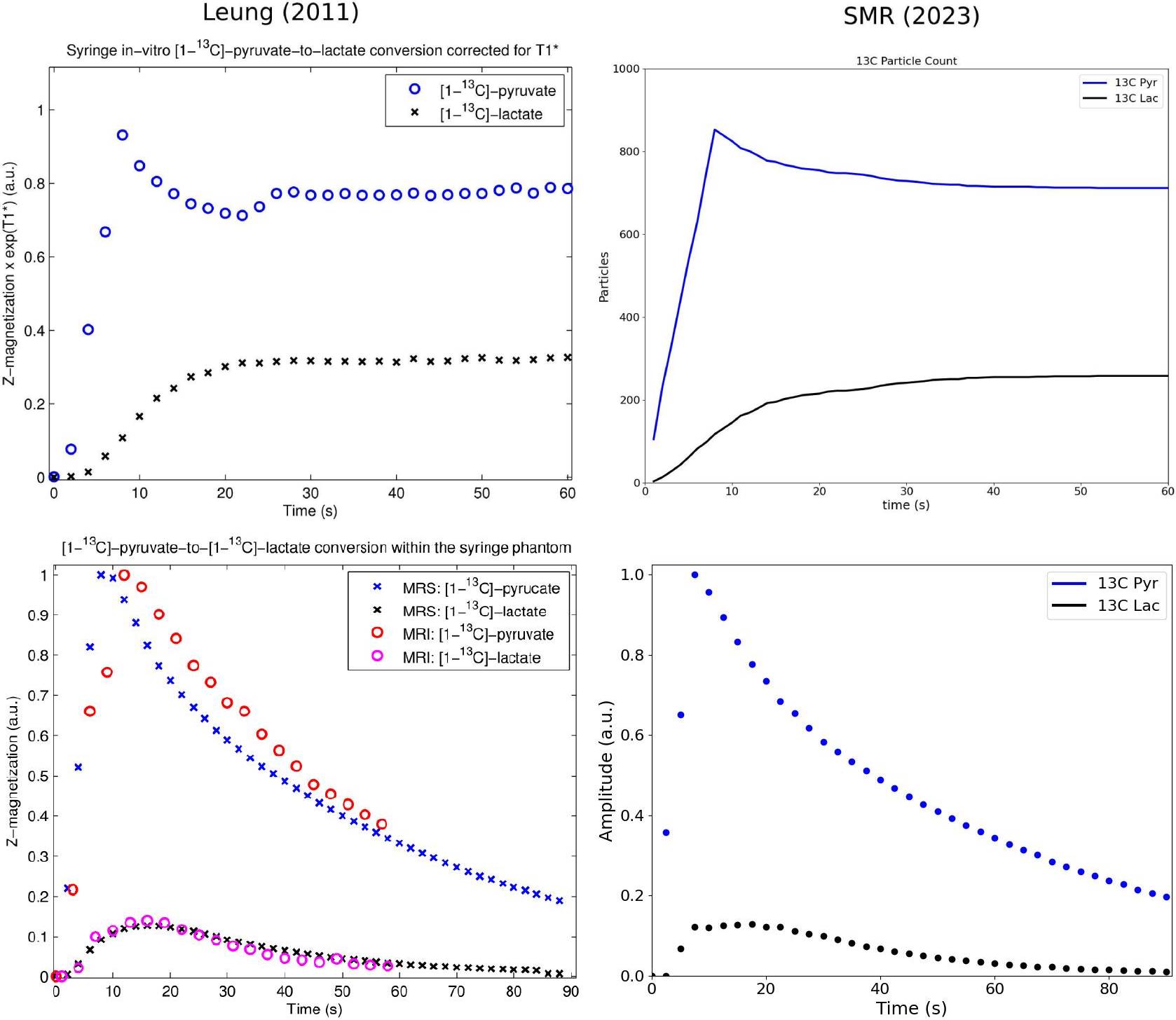
Simulations were conducted using the particle-based biochemical simulator Smoldyn7 with the MR module described previously.8 The core model was a 10 micron cubic volume containing 12C and 13C metabolites and LDH complexes bound with NADH or NAD+. Diffusion coefficients reference values were: 1120 μm2/s pyruvate,9 1000 μm2/s lactate,9,10 and 49.9 μm2/s LDH.11 Both 12C and 13C metabolites could react with enzyme-coenzyme complexes (LDH-NADH, LDH-NAD+), with an initial NADH:NAD+ ratio of 10. Reaction rate constants were fixed for a time-to-equilibrium under 60 s. Initial particle populations were 100 12C-pyruvate, 1,000 13C-pyruvate, and 12C-lactate from 100 to 10,000, and no 13C-lactate. Particle populations were tracked over 60 s simulation time and counts of reacting metabolites were recorded at the start (0-1 s) and end (55-60 s).MR spectroscopic results from an in vitro isotope exchange experiment12 were replicated using this model. In the reference experiment, hyperpolarized 13C-pyruvate was injected over 10 s into a syringe containing 40 mM 12C-lactate, 20 mM NAD+, 10 mM NADH, and LDH-5. In silico, a zeroth-order reaction was added to instantiate pyruvate particles over 10 s, followed by another 80 s simulation time. Spectroscopic acquisitions of the phantom volume were taken over 90 s using a 2.5° flip angle pulse with 5000 Hz bandwidth, and a matching sequence was used to obtain simulated spectroscopic data.
Results & Discussion
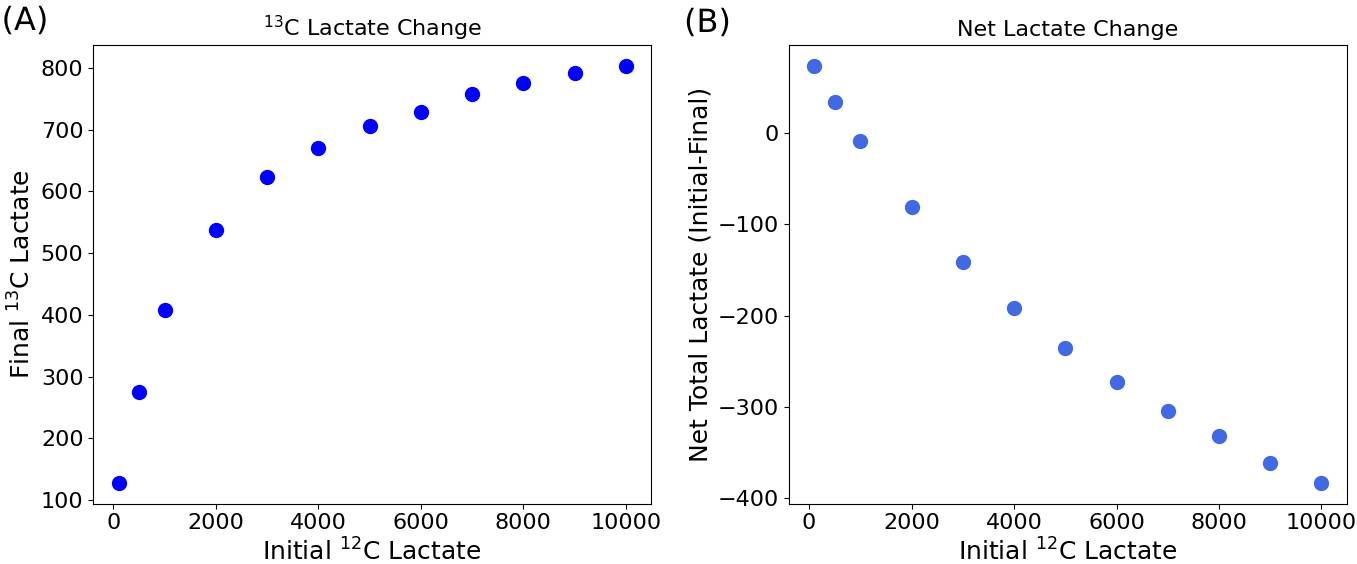
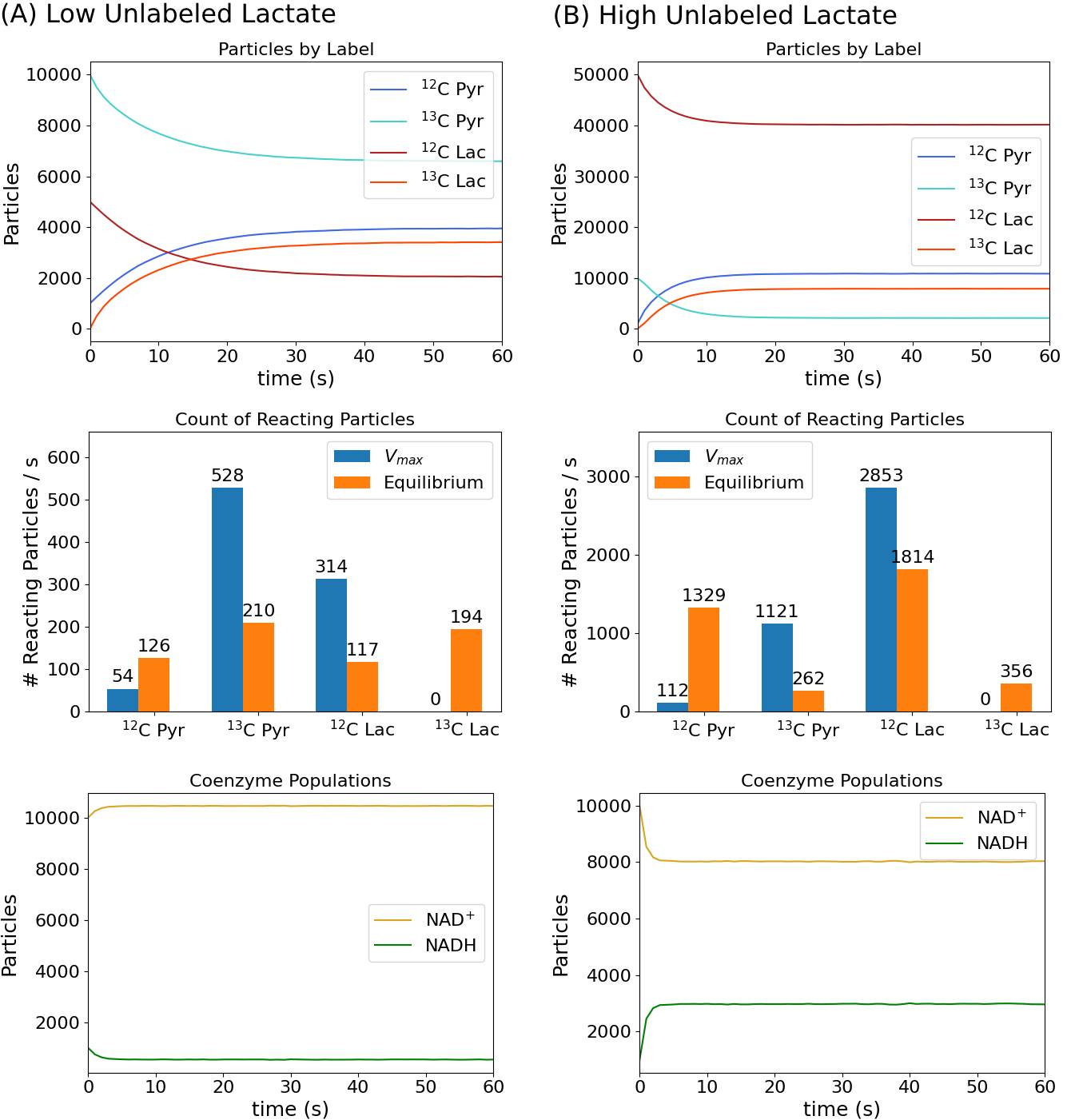
Net production of [1-13C]lactate and change in total lactate with variable initial populations of unlabeled lactate are plotted in Figure 2. As expected, higher levels of 12C-lactate increase production of [1-13C]lactate, with gradually diminishing effects from marginal increases in unlabeled lactate. Simultaneously, the net change in total lactate has a decreasing trend. Figure 3 provides further insight into this phenomenon. In the lower lactate case, with [1-13C]pyruvate as the most populated substrate, net lactate production occurs. In the higher lactate case, the number of reactions increases due to the increased overall particle population, so [1-13C]lactate production, exchange, and equilibration of the system are all faster; however, the high population of 12C-lactate also means that unlabeled metabolites now participate in a higher proportion of reactions relative to labeled metabolites. Under such conditions, the total lactate population goes down, since the balance of rate constants favors an equilibrium with a lower proportion of lactate than in the initial populations. Increased [1-13C]lactate is thus not straightforwardly correlated with overall metabolic production of lactate. Because the model implements isotope exchange as a stochastic particle simulation, it is capable of replicating non-linear dynamics which are difficult to describe with first-order kinetic rate equations alone, such as when kinetics are disrupted by depletion of the coenzyme pool. Accurate simulation of such a case from a phantom experiment is plotted in Figure 4: particle population counts (top right) reproduce the in vitro time course (top left) after the latter was corrected for loss of magnetization due to flip-down magnetization and T1 decay of hyperpolarized substrates. Pyruvate and lactate peak amplitudes from MR spectroscopic measurements also align closely with simulated values using an equivalent pulse sequence (Figure 4, bottom row).Conclusions
This model is a realistic and versatile in silico implementation of metabolic isotope exchange, a process which is critical to understanding the physiological relevance of HP 13C MR results. The model generated results consistent with an in vitro isotope exchange experiment that deviated from simple first-order kinetic rate equations, while providing additional information through particle-scale data.Acknowledgements
No acknowledgement found.References
(1) Day, S. E.; Kettunen, M. I.; Gallagher, F. A.; Hu, D.-E.; Lerche, M.; Wolber, J.; Golman, K.; Ardenkjaer-Larsen, J. H.; Brindle, K. M. Detecting Tumor Response to Treatment Using Hyperpolarized 13C Magnetic Resonance Imaging and Spectroscopy. Nat. Med. 2007, 13 (11), 1382–1387. https://doi.org/10.1038/nm1650.
(2) Brooks, G. A. Lactate as a Fulcrum of Metabolism. Redox Biol. 2020, 35, 101454. https://doi.org/10.1016/j.redox.2020.101454.
(3) Rabinowitz, J. D.; Enerbäck, S. Lactate: The Ugly Duckling of Energy Metabolism. Nat. Metab. 2020, 2 (7), 566–571. https://doi.org/10.1038/s42255-020-0243-4.
(4) Li, X.; Yang, Y.; Zhang, B.; Lin, X.; Fu, X.; An, Y.; Zou, Y.; Wang, J.-X.; Wang, Z.; Yu, T. Lactate Metabolism in Human Health and Disease. Signal Transduct. Target. Ther. 2022, 7 (1), 1–22. https://doi.org/10.1038/s41392-022-01151-3.
(5) Brindle, K. M. Imaging Metabolism with Hyperpolarized 13C-Labeled Cell Substrates. J. Am. Chem. Soc. 2015, 137 (20), 6418–6427. https://doi.org/10.1021/jacs.5b03300.
(6) Wang, Z. J.; Ohliger, M. A.; Larson, P. E. Z.; Gordon, J. W.; Bok, R. A.; Slater, J.; Villanueva-Meyer, J. E.; Hess, C. P.; Kurhanewicz, J.; Vigneron, D. B. Hyperpolarized 13C MRI: State of the Art and Future Directions. Radiology 2019, 291 (2), 273–284. https://doi.org/10.1148/radiol.2019182391.
(7) Andrews, S. S. Smoldyn: Particle-Based Simulation with Rule-Based Modeling, Improved Molecular Interaction and a Library Interface. Bioinformatics 2017, 33 (5), 710–717. https://doi.org/10.1093/bioinformatics/btw700.
(8) (ISMRM 2022) Novel particle-based spatial stochastic Bloch simulation applied to LDH-mediated pyruvate conversion. https://archive.ismrm.org/2022/1425.html (accessed 2022-10-04).
(9) Koelsch, B. L.; Keshari, K. R.; Peeters, T. H.; Larson, P. E. Z.; Wilson, D. M.; Kurhanewicz, J. Diffusion MR of Hyperpolarized 13C Molecules in Solution. The Analyst 2013, 138 (4), 1011–1014. https://doi.org/10.1039/c2an36715g.
(10) Ribeiro, A. C. F.; Lobo, V. M. M.; Leaist, D. G.; Natividade, J. J. S.; Veríssimo, L. P.; Barros, M. C. F.; Cabral, A. M. T. D. P. V. Binary Diffusion Coefficients for Aqueous Solutions of Lactic Acid. J. Solut. Chem. 2005, 34 (9), 1009–1016. https://doi.org/10.1007/s10953-005-6987-3.
(11) Squire, P. G.; Himmel, M. E. Hydrodynamics and Protein Hydration. Arch. Biochem. Biophys. 1979, 196 (1), 165–177. https://doi.org/10.1016/0003-9861(79)90563-0.
(12) Leung, K. K.-C. Dynamic Interleaved Imaging of Pyruvate Metabolism with Hyperpolarized 13C. Thesis, 2011. https://tspace.library.utoronto.ca/handle/1807/27311 (accessed 2023-11-03).
Figures