4537
EPR imaging of oxygen consumption driven by NQO1-activated compounds in FH-deficient renal tumors1Urologic Oncology Branch, National Cancer Institute, National Institution of Health, Bethesda, MD, United States, 2National Institute of Neurological Disorders and Stroke, National Institution of Health, Bethesda, MD, United States, 3Radiation Biology Branch, National Cancer Institute, National Institution of Health, Bethesda, MD, United States
Synopsis
Keywords: Preclinical Image Analysis, Electron Paramagnetic Resonance
Motivation: Renal tumors in patients affected by Hereditary Leiomyomatosis and Renal Cell Cancer (HLRCC) are aggressive and metastasize early. The high expression of NAD(P)H quinone oxidoreductase 1 (NQO1) in HLRCC could be a therapeutic target.
Goal(s): Our goal was to determine if NQO1-activated substrates can be used as a therapeutic approach for HLRCC renal tumors.
Approach: In vitro experiments and EPR, DCE, and photoacoustic imaging were performed to study the effect of NQO1-activated substrates on oxygen levels and tumor growth in HLRCC.
Results: Our study demonstrated by EPR imaging that oxygen consumption is induced by NQO1-activated substrates, resulting in ROS production and tumor cytotoxicity.
Impact: We found that NQO1-activated substrates induced non-mitochondrial oxygen consumption in FH-deficient renal tumor cells, triggering oxidative stress-induced cancer cell death. Our results suggest a promising path for NQO1-targeted therapy in HLRCC, necessitating further research and treatment development.
Introduction
Hereditary renal cell carcinoma is a disease with different pathologies, clinical courses, and therapeutic treatments1. Among them, HLRCC is characterized by FH gene mutations in the TCA cycle associated with mitochondrial DNA abnormalities, resulting in a pronounced metabolic shift toward glycolysis1,2. Additionally, FH mutations lead to fumarate accumulation, triggering upstream NRF2 signaling and significantly elevating the expression of NQO1 compared to normal renal tissue1,3. While quinones such as β-lapachone and isobutyl-deoxynyboquinone (IB-DNQ), several NQO1 substrates with potential anticancer effects have been reported, their therapeutic applications in renal cell cancer, particularly HLRCC, have been underexplored4. Therefore, in this study, we investigated the NQO1-dependent cancer therapeutic effects of the target drugs in in vitro studies and their effects on HLRCC tumor oxygenation levels using in vivo imaging techniques with EPR, DCE-MRI, and photoacoustic imaging.Methods
UOK262 and UOK365 cell line were established from FH-deficient renal tumors and the UOK342 cell line was derived from type 1 papillary RCC patient who had surgery at the NIH Clinical Center. NQO1 KO UOK262 cell lines were established by CRISPR-Cas9 system. ROS production by quinone treatments were measured by DCFDA fluorescent probe. Cytotoxicity of quinones were determined using live-cell analysis system (Incucyte®). NOD scid gamma (NSG) mice were subcutaneously injected with these cell lines in the hind leg. Quinones were administered to the mice by intravenous tail vein injection. In vitro oxygen consumption rates of these cell lines were measured using an XFe Extracellular Flux Analyzer from Seahorse Biosciences. Parallel coil resonators tuned to 300 MHz were used for EPR imaging. OX063 (1.125 mmol/kg bolus) was injected intravenously to a mouse through a tail vein cannula. The free induction decay (FID) signals were collected following the radiofrequency excitation pulses (65 ns) with a nested looping of the x, y, and z gradients, and each time point in the FID underwent phase modulation, enabling 3D spatial encoding (Repetition time: 8.0 μs, Number of averages: 4,000). Following EPR imaging, anatomic T2-weighted MR images were collected with a 1T scanner. DCE-MRI studies were performed on a 3 T scanner (Bruker BioSpec 3T). T1-weighted fast low-angle shot (FLASH) images were obtained with TR = 117.2 ms; TE = 6 ms; flip angle = 30˚; two slices; 28 x 28 mm resolution; 15-second acquisition time per image; and 45 repetitions. Gd-DTPA solution (4 mL/g of body weight of 50 mmol/L Gd-DTPA) was injected through a tail vein cannula 1 minute after the start of the dynamic FLASH sequence. To determine the local concentrations of Gd-DTPA, T1 maps were calculated from three sets of Rapid Imaging with Refocused Echoes (RARE) images obtained with TR = 320, 400, 600, 1,000, 2,000, and 3,000 ms, with the acquisitions being made before running the FLASH sequence. Photoacoustic imaging studies were performed with Vevo LAZR-X. Tumor images were observed with the MX250 probe and the tumor oxygen saturation was measured in PA-mode.Results
Our study revealed that HLRCC cells exhibit minimal oxygen consumption in vitro, and in vivo experiments demonstrated higher oxygen levels within HLRCC tumors (26.2 mmHg; UOK262, 24.9 mmHg; UOK365) compared to type 1 papillary renal cancer cell line UOK342 (16.4 mmHg), which showed relatively robust respiration in vitro (Fig 1). Introduction of quinones to HLRCC cells resulted in stimulation of non-mitochondrial oxygen consumption and the production of reactive oxygen species, leading to cytotoxicity (Fig 2A, B). In addition, NQO1 KO UOK262 cells were less sensitive to quinones compared to control UOK262 cells (Fig 2C, D). Furthermore, utilizing EPR and photoacoustic in vivo imaging, we observed a reduction in oxygen levels within HLRCC tumors following administration of the quinones (Fig 3).Discussion
Intratumor oxygen levels were found to be higher in tumors with low mitochondrial oxygen consumption. We propose that β-lapachone and IB-DNQ promote non-mitochondrial oxygen consumption in an NQO1-dependent manner and induces oxidative stress-induced cell death in FH-deficient renal tumor xenografts. Furthermore, in two different oxygen imaging experiments using HLRCC tumor mouse models, quinone treatment markedly reduced intratumoral oxygen levels, suggesting that these imaging results may reflect therapeutic efficacy.Conclusion
In conclusion, our study highlights the promise of targeting NQO1 in HLRCC tumors, which rely heavily on glycolysis and exhibit high intratumoral oxygen levels compared to other renal cancer types. This novel therapeutic approach may extend beyond HLRCC to cancers with elevated NQO1 expression in various organs. Moreover, the effectiveness of this treatment strategy could be predicted using oxygen imaging techniques such as EPR and photoacoustic imaging. Further research is warranted to explore these promising avenues for cancer therapy.Acknowledgements
We are deeply grateful to members of Radiation Biology Branch, National Institute of Health for their skillful technical assistance.References
1. Linehan WM, Schmidt LS, Crooks DR, et al. The metabolic basis of kidney cancer. Cancer Discov. 2019;9;1006 – 1021.
2. Crooks DR, Maio N, Lang M, et al. Mitochondrial DNA alteration underlies an irreversible shift to aerobic glycolysis in fumarate hydratase-deficient renal cancer. Sci Signal. 2021;14;eabc4436.
3. Sourbier C, Ricketts CJ, Matsumoto S, et al. Targeting ABL1-mediated oxidative stress adaptation in fumarate hydratase-deficient cancer. Cancer Cell. 2014;26;840 – 850.
4. Parkinson EI and Hergenrother PJ. Deoxynyboquinones as NQO1-activated cancer therapeutics. Acc Chem Res. 2015;48;2715 – 2723.
Figures
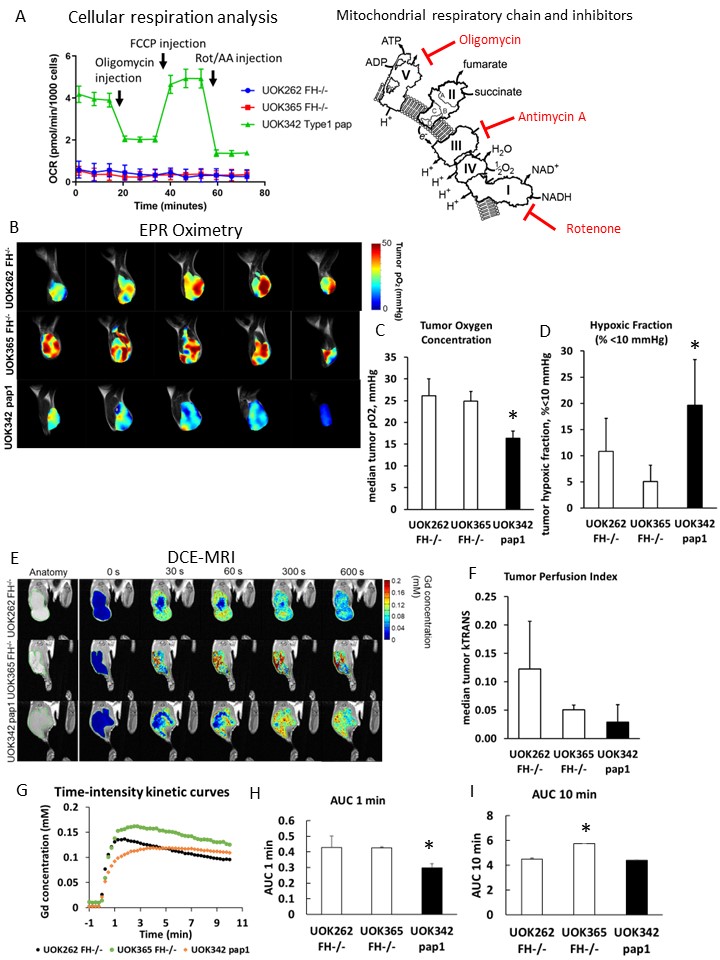
FH-deficient xenografts exhibit high tumor oxygen levels that are independent of tumor perfusion level.
(A) Mito stress test of RCC cell lines (1 µM oligomycin, 1 µM FCCP and 0.5 1 µM rotenone/antimycin A). (B) Representative EPR images of RCC xenografts in NSG mice. (C and D) Difference of pO2 (C) and HF10 (D) between RCC xenografts. (E) Representative images of DCE-MRI, (F) Ktrans median, (G) time-intensity kinetic curves, (H) AUC 1 min, and (I) AUC 10 min of each RCC xenograft. Data are expressed as mean ± SD. Statistical analysis by Welch’s t-test (*p < 0.05).
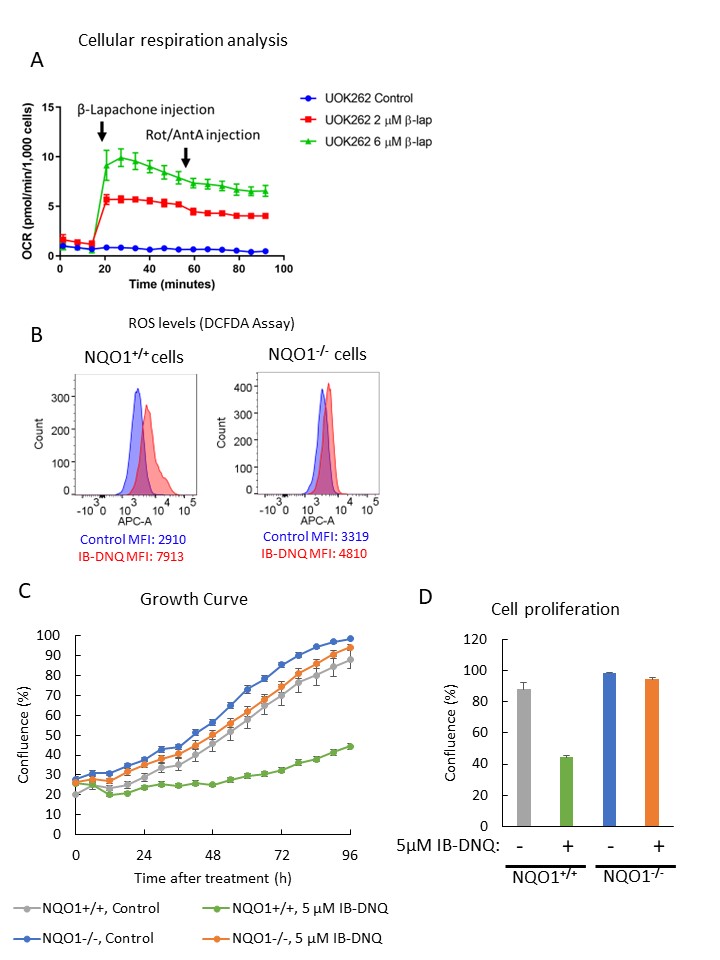
NQO1-activated quinones stimulated non-mitochondrial respiration and oxidative stress in FH-deficient tumor cells in an NQO1-dependent manner.
(A) Measurement of oxygen consumption rate stimulated by β-lapachone treatment in FH-deficient UOK262 cells. (B) Measurement of intracellular ROS induced by IB-DNQ treatment by DCFDA assay on CRISPR-edited UOK262 cells. (C and D) Cytotoxicity assay of IB-DNQ on CRISPR-edited UOK262 cells. Data are expressed as mean ± SD.
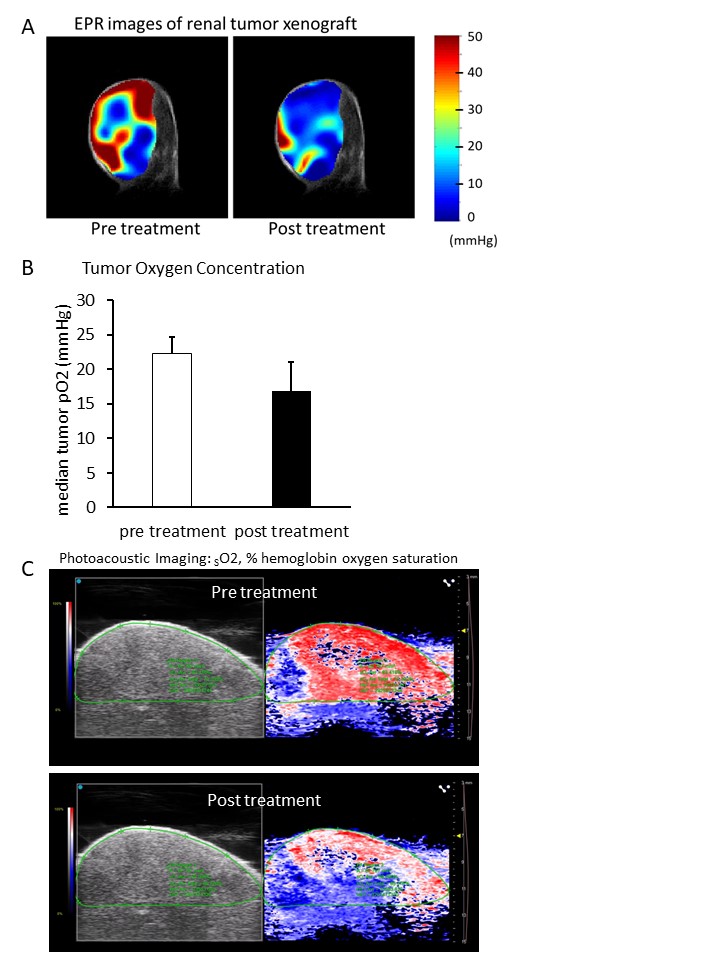
Stimulation of non-mitochondrial respiration in vivo with IB-DNQ therapy resulted in an acute decrease in tumor oxygen levels.
(A) EPR oximetry imaging of UOK262 xenografted NSG mice treated with 20 mg/kg β-lapachone. (B) Median tumor pO2 calculated by EPR oximetry imaging. Data are expressed as mean ± SEM. (C) Photoacoustic oximetry imaging on UOK262 xenografted NSG mice with 5 mg/kg IB-DNQ treatment.