4532
Enhancement of Radiation Therapy Response in Prostate Cancer by Metabolic Modulation1University of Pennsylvania, Philadelphia, PA, United States, 2Advanced Imaging Research, Inc., Cleveland, OH, United States, 3Medical College of Wisconsin, Milwaukee, WI, United States
Synopsis
Keywords: Other Preclinical, Molecular Imaging, Prostate Cancer Models, Lonidamine, mito-Lonidamine, 1H and 31P MRS, Seahorse, oxygen consumption rate, pH
Motivation: When prostate cancer is treated with external beam radiation therapy (RT) with doses up to 78 Gy, gastrointestinal and genitourinary toxicities are often observed.
Goal(s): Tumor sensitization by mito-lonidamine (mito-LND) will lower RT doses reducing the risk of adverse effects.
Approach: The effects were assessed in vitro and in vivo in prostate cancer models using Seahorse, 1H and 31P MRS respectively.
Results: Our findings showed a sustained and tumor-selective decrease in intracellular pH, bioenergetics, oxygen consumption rate and lactate. Selective tumor acidification, deenergization and oxygenation induced by mito-LND may improve the radiation response in prostate cancer.
Impact: Exploiting the modulation of tumor metabolism and microenvironment for improving therapeutic efficacy of radiation therapy (RT) in early stage prostate cancer will lead to improved outcomes in prostate cancer patients.
Introduction
Prostate cancer is the most common cancer and the second-leading cause of cancer death in men in the United States.1 When detected early, prostate cancer is usually treated with local therapy, mainly surgery and/or radiation treatment. Early prostate cancer often presents as a multi-focal disease and standard dose escalated radiation therapy (RT) in the range of 78 Gy is associated with gastrointestinal (GI) & genitourinary (GU) toxicities. We hypothesize that tumor sensitization by mito-lonidamine (mito-LND) will enable the use of lower RT doses reducing risk of side effects in androgen dependent and androgen independent prostate cancer xenografts. Using mitochondria-targeted mito-LND could markedly enhance its potency and/or efficacy to cancer inhibition using RT.2 Mito-LND, as a tumor-selective inhibitor of oxidative phosphorylation, inhibits ATP production, oxygen consumption, GSH and induces reactive oxygen species to selectively sensitize to RT and is 100-fold more effective compared to lonidamine (LND).Methods
PC3, androgen independent and CWR22Rv1, androgen dependent prostate cancer cells were grown in RPMI 1640 medium supplemented with 10% fetal bovine serum, 2mM L-glutamine, and 1% penicillin-streptomycin. In vitro studies: In vitro oxygen consumption and extracellular acidification rates (OCR and ECAR) were determined using the Seahorse XF-96 Extracellular Flux Analyzer with and without LND (200 µM) and mito-LND (2 µM) treatment. Glucose and lactate concentrations were measured using YSI 2300 STAT Plus Glucose & Lactate Analyzer under similar conditions. In vivo studies: 7×106 PC3 cells were inoculated subcutaneously in each mouse (n=5) as a 0.1 mL suspension. MR experiments were performed on a 9.4 T/31 cm horizontal bore Bruker spectrometer. 1H and 31P MRS experiments were performed after positioning the s.c. tumor in a dual-frequency slotted-tube resonator as described elsewhere;3 the intracellular pH (pHi; n=5), extracellular pH (pHe; n=5), bioenergetics (βNTP/Pi; n=5), and lactate (n=5) concentration were measured before and 14 days post LND and mito-LND with dose of 5.4 mg/kg oral; once daily, up to 14 days). Physiological monitoring was maintained during the experiment. Procedures for data acquisition, post processing and parameter estimation were performed as previously described.3 Small Animal Radiation Research Platform (SARRP) was used to irradiate the PC3 prostate cancer cells to assess the effect of LND and mito-LND on RT. Various radiation doses were used to determine the dose response curve using clonogenic assay. Based on the clonogenic dose response curve, 2 Gy dose was selected for sufficient dynamic range to demonstrate increased response with LND and mito-LND. Radiation dose was applied after 40 min. of LND and mito-LND administration to achieve significant acidification and deenergization. The data on pHi, pHe, bioenergetics and lactate at various time points following LND and mito-LND administration were compared by t-test analysis.Results
Our metabolic studies of extracellular glucose and lactate using the YSI 2300 STAT PLUS Glucose Lactate Analyzer have shown that mito-LND significantly increases glucose consumption and lactate production in PC3 and CWR22Rv1 cells (Figure 1). Mito-LND also significantly decreased the oxygen consumption rate of PC3 and CWR22Rv1 cells, resulting in lower rates of oxidative phosphorylation and the shifting of cells to a more glycolytic energy phenotype (Figure 2). Clonogenic assays carried out on PC3 cells treated with mito-LND in combination with 2 Gy radiation have shown that mito-LND significantly enhanced the effect of radiation therapy (Figure 3). 1H MRS studies on PC3 mouse tumor xenografts showed a significant decrease in total lactate levels with chronic LND and mito-LND treatment over 14 days, with mito-LND being more potent (Figure 4). Furthermore, 31P MRS studies showed that chronic treatment with mito-LND significantly decreased intracellular pH and extracellular pH in mouse tumors compared to control and LND and caused a significant decrease in tumor bioenergetics (Figure 5).Discussion
Prostate cancer cells treated with mito-LND increased rates of glycolysis to compensate for the inhibition of oxidative phosphorylation through the blocking of complex I and II of the electron transport chain. This is demonstrated by our in vitro YSI and Seahorse experiments. In the short-term, cells treated with mito-LND upregulated glycolysis to compensate for the loss of oxidative phosphorylation. In the long-term, chronic treatment of prostate cancer cells with mito-LND caused a significant overall decrease in cellular energy, which was demonstrated by our 1H and 31P MRS experiments on mouse tumor xenografts, and loss of viability, making mito-LND a potent anti-cancer drug.Conclusions
Our data show that mito-LND significantly inhibits mitochondrial respiration, more potently than LND. Mito-LND could increase intracellular oxygen concentrations and potentiate the effect of radiation therapy, leading to effective radiation therapy with lower toxicity and potentially fewer detrimental side effects in prostate cancer patients.Acknowledgements
This work was supported in part by grants from National Cancer Institute 1R01CA250102, 1R01CA228457, 1R01CA268601 and Institute for Translational Medicine and Therapeutics (ITMAT) of the University of Pennsylvania.References
1. Siegel RL, Miller KD, Wagle NS, et al. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17-48.
2. Cheng G, Zhang Q, Pan J, et al. Targeting lonidamine to mitochondria mitigates lung tumorigenesis and brain metastasis. Nat Commun. 2019;10(1):2205.
3. Nath K, Nelson DS, Ho AM, et al. 31P and 1H MRS of DB-1 melanoma xenografts: lonidamine selectively decreases tumor intracellular pH and energy status and sensitizes tumors to melphalan. NMR Biomed. 2013;26(1):98-105.
Figures
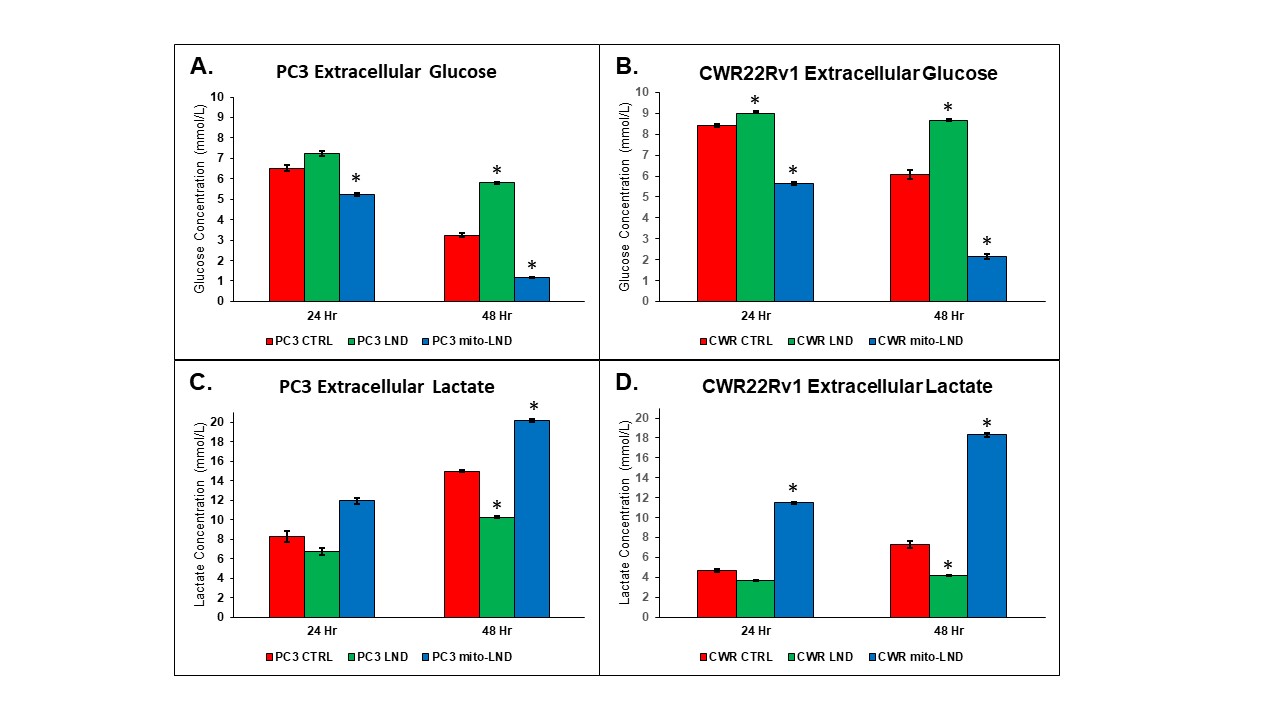
Figure 1. Effect of LND and mito-LND on growth of PC3 and CWR22Rv1 cells
Effect of LND and mito-LND on PC3 and CWR22Rv1 prostate cancer cells. Cells were treated with 200 µM LND and 2 µM mito-LND and examined at the 24- and 48-hour time points using extracellular glucose, (A) and (B), and lactate concentrations, (C) and (D), gathered with the YSI 2300 STAT PLUS Biochemical Analyzer. All experiments were performed at n=3 and data are represented as the mean ± standard error mean with statistical significance of p < 0.01 indicated by the asterisks.
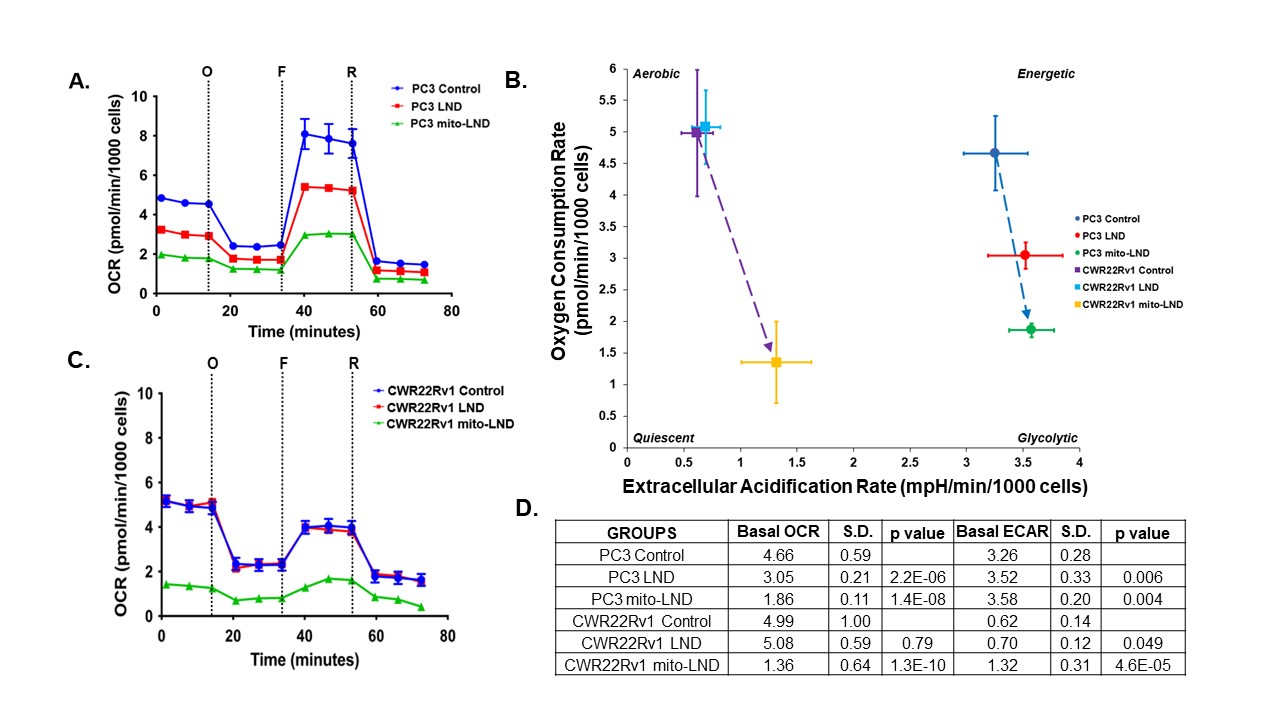
Figure 2. Seahorse Mitochondrial Function Assay
Effect of 200 µM LND and 2 µM mito-LND on mitochondrial function of PC3 and CWR22Rv1 cells. Mito Stress Test Assay was carried out using the following reagents: Oligomycin, “O”, FCCP, “F”, and Rotenone/Antimycin-A, “R”. (A) OCR of PC3 cells. (B) Energy map showing metabolic shifts and changes in cell energy phenotype. (C) OCR of CWR22Rv1 cells. (D) Quantified OCR and ECAR values comparing control to treated groups. Data were gathered on Seahorse XFe96 Analyzer. Cells were treated for 24 hours with sample size of n=14 per group.
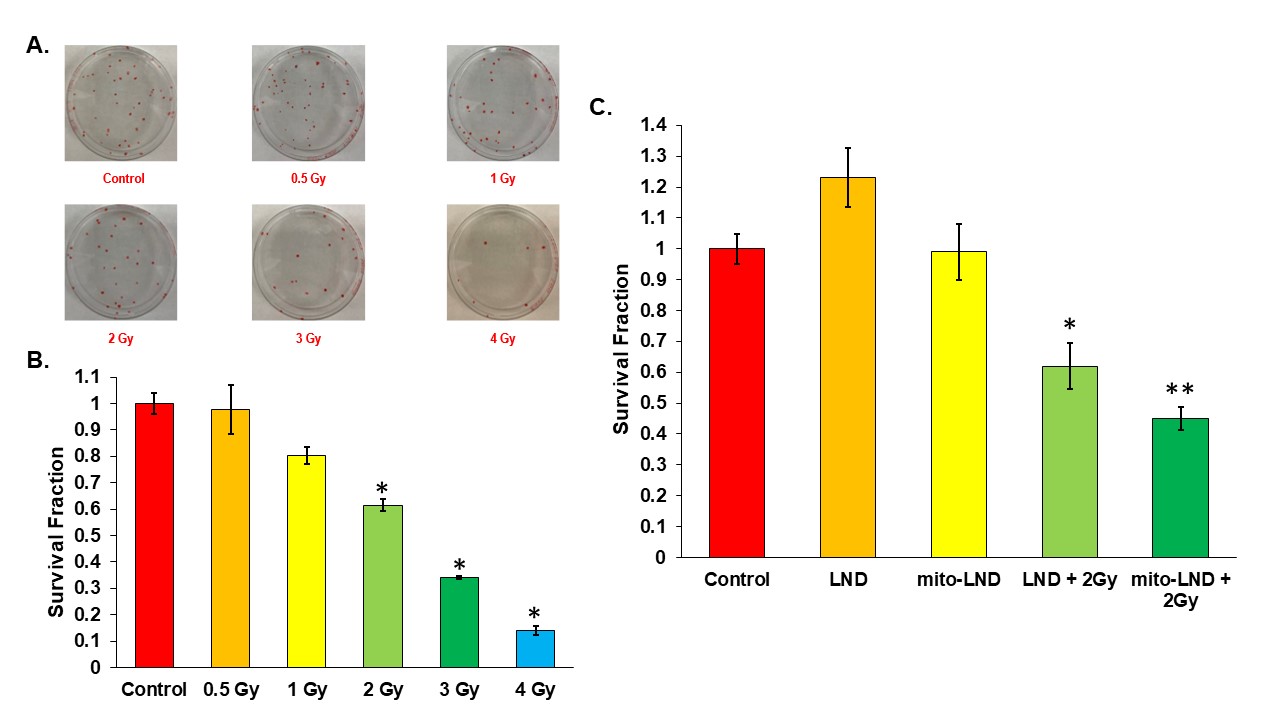
Figure 3. Radiation Enhancement with LND and mito-LND
(A) Plates showing counted colonies of a clonogenic assay carried out on PC3 cells at various doses of radiation. (B) Figure shows the survival fraction of PC3 cells at various doses of radiation. (C) Figure shows the survival fraction of PC3 cells treated with 200 µM LND and 2 µM mito-LND in combination with 2 Gy radiation. All experiments were performed in triplicate and the data are represented as mean ± standard error mean with statistical significance of p < 0.01 indicated by the asterisks.
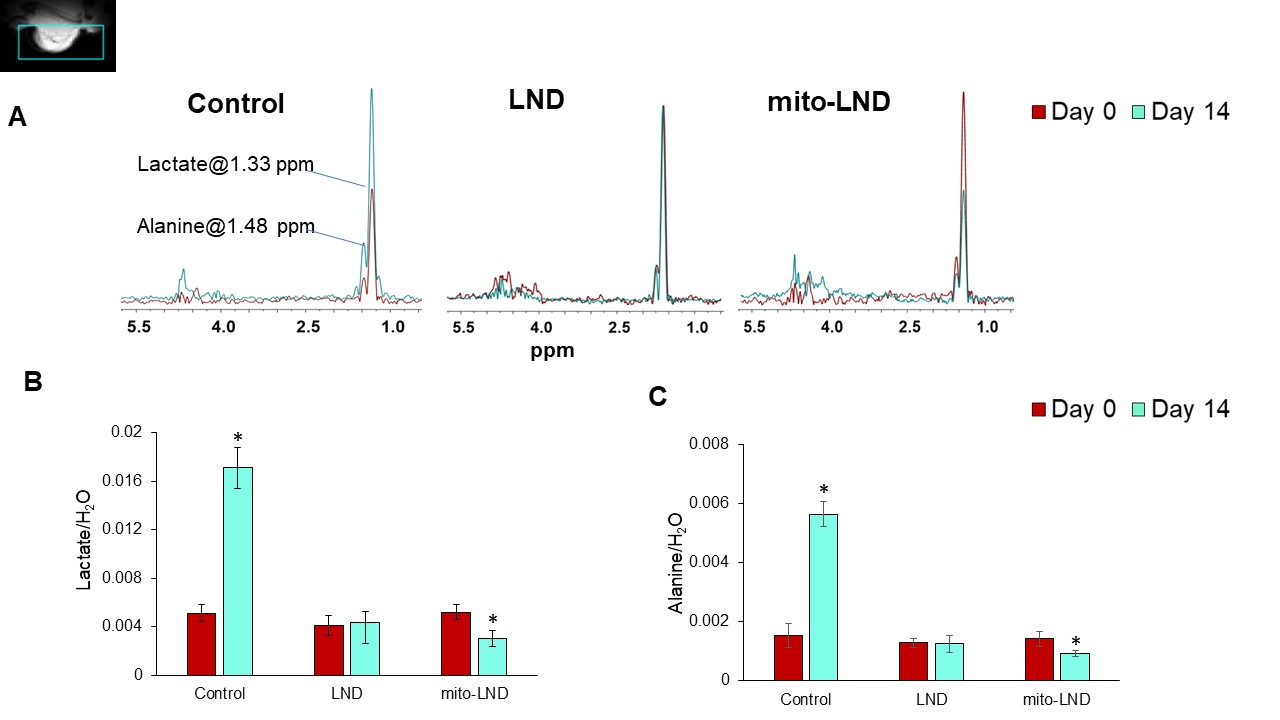
Figure 4. In Vivo 1H MRS
(A) Representative overlaid 1H MRS with HDMD-Sel-MQC pulse sequence spectra in each group. The spectral peak area of (B) lactate, (C) alanine signals normalized to the water signal measured by 1H MRS with HDMD-Sel-MQC transfer pulse sequence in PC3 prostate cancer xenografts (n=5) after treatment with LND and mito-LND with the dose of 5.4 mg/kg, orally, once daily up to 14 days. We have also included vehicle treated controls.
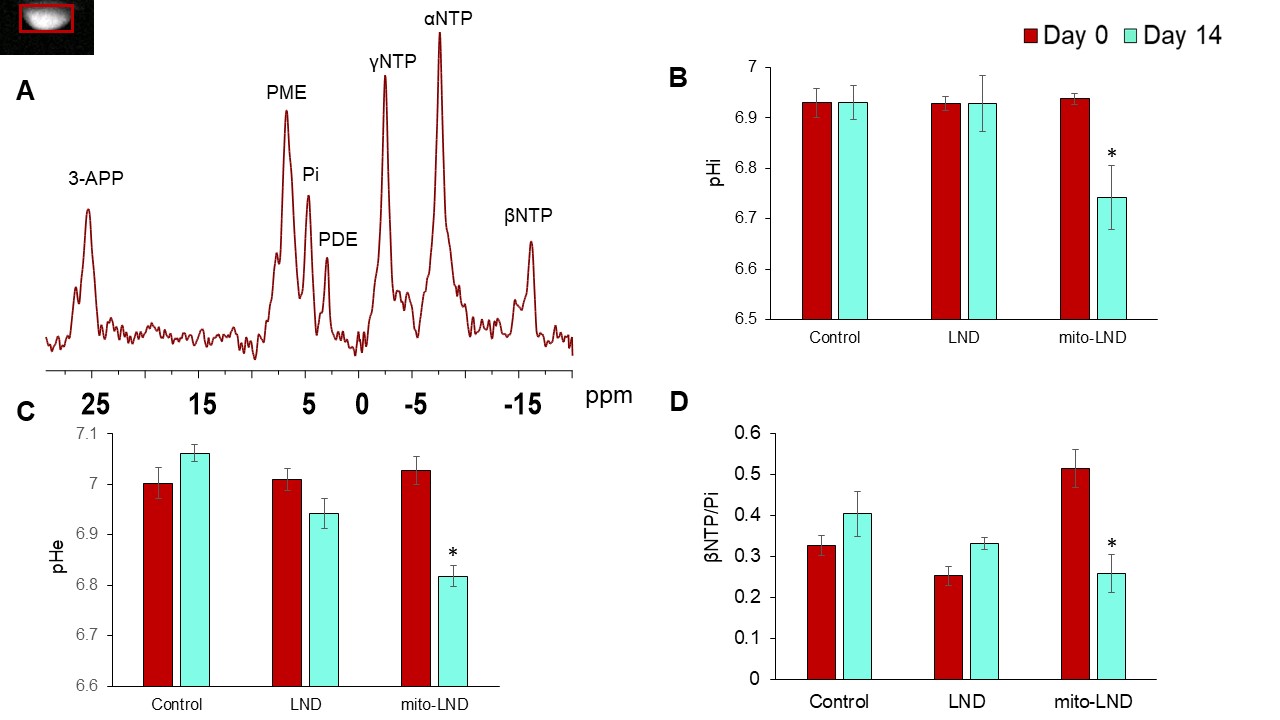
Figure 5. In Vivo 31P MRS
(A) Representative in vivo localized 31P MRS of PC3 prostate cancer xenografts. (B) Intracellular pH (pHi) (C) Extracellular pH (pHe) and (D) Bioenergetics (βNTP/Pi) measured using 31P MRS in PC3 prostate cancer xenografts (n=5) after treatment with LND and mito-LND with the dose of 5.4 mg/kg, orally, once daily up to 14 days. We have also included vehicle treated controls.