4472
Superior HyperCEST Performance of Membrane-Anchored Xenon Hosts in Nanocarriers with Variable Membrane Fluidity1Translational Molecular Imaging, German Cancer Research Center, Heidelberg, Germany, 2Department for Physics and Astronomy, Ruprecht Karls University Heidelberg, Heidelberg, Germany, 3German Cancer Consortium (DKTK), Heidelberg, Germany
Synopsis
Keywords: Contrast Agents, Contrast Agent
Motivation: The efficiency of CEST agents for hyperpolarized 129Xe depends on the exchange rate of Xe in/out of tailored host structures. For liposomal designs, this may be influenced by the phospholipid membrane fluidity.
Goal(s): This study investigates how cholesterol, which is often added for liposome stability, impacts the HyperCEST performance.
Approach: We compared the changes in CEST buildup from liposomes with variable cholesterol content and either membrane-anchored (i.e., lipopeptide-based) or freely diffusing Xe host.
Results: The HyperCEST efficiency for membrane-anchored Xe hosts is much less sensitive to membrane stiffening than for unbound hosts. Lipopeptide-based HyperCEST agents are thus a powerful approach for biosensor design.
Impact: The HyperCEST performance of membrane-anchored xenon hosts in liposomal nanocarriers shows reduced susceptibility to membrane stiffening compared to non-functionalized hosts. These less susceptible lipopeptide-based hosts are thus the preferred approach for future in vivo applications.
Introduction
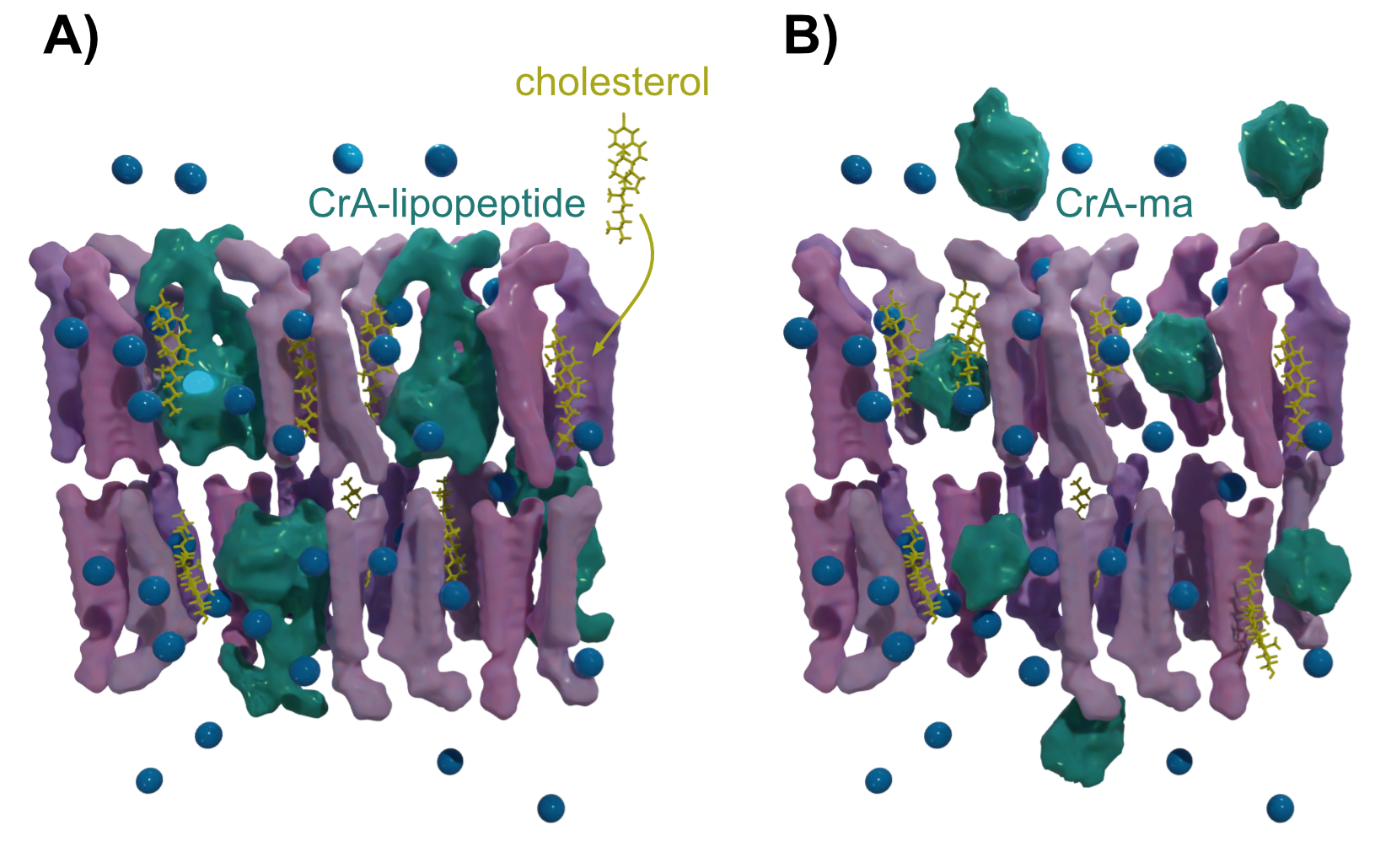
HyperCEST with xenon biosensors is an emerging technique that has shown unprecedented sensitivity for targeted contrast agents in vitro. However, it still lacks efficient reporters that will work under in vivo conditions. We have recently introduced a liposomal approach[1] in which high numbers of lipopeptide-anchored Xe hosts are embedded into phospholipid membranes of nanocarriers (Figure 1A). The membranes of liposomal carriers often contain cholesterol for stabilization. This reduces the membrane fluidity and can thus impact the Xe exchange rate and, eventually, reduce the HyperCEST performance. This effect has been reported to be quite pronounced for freely diffusing Xe hosts[2] (Figure 1B) and would thus cause unwanted loss in the efficiency of such HyperCEST agents. We therefore investigated in this study how different cholesterol content impacts the HyperCEST performance and compared the results from membrane-anchored and freely diffusing hosts.Methods
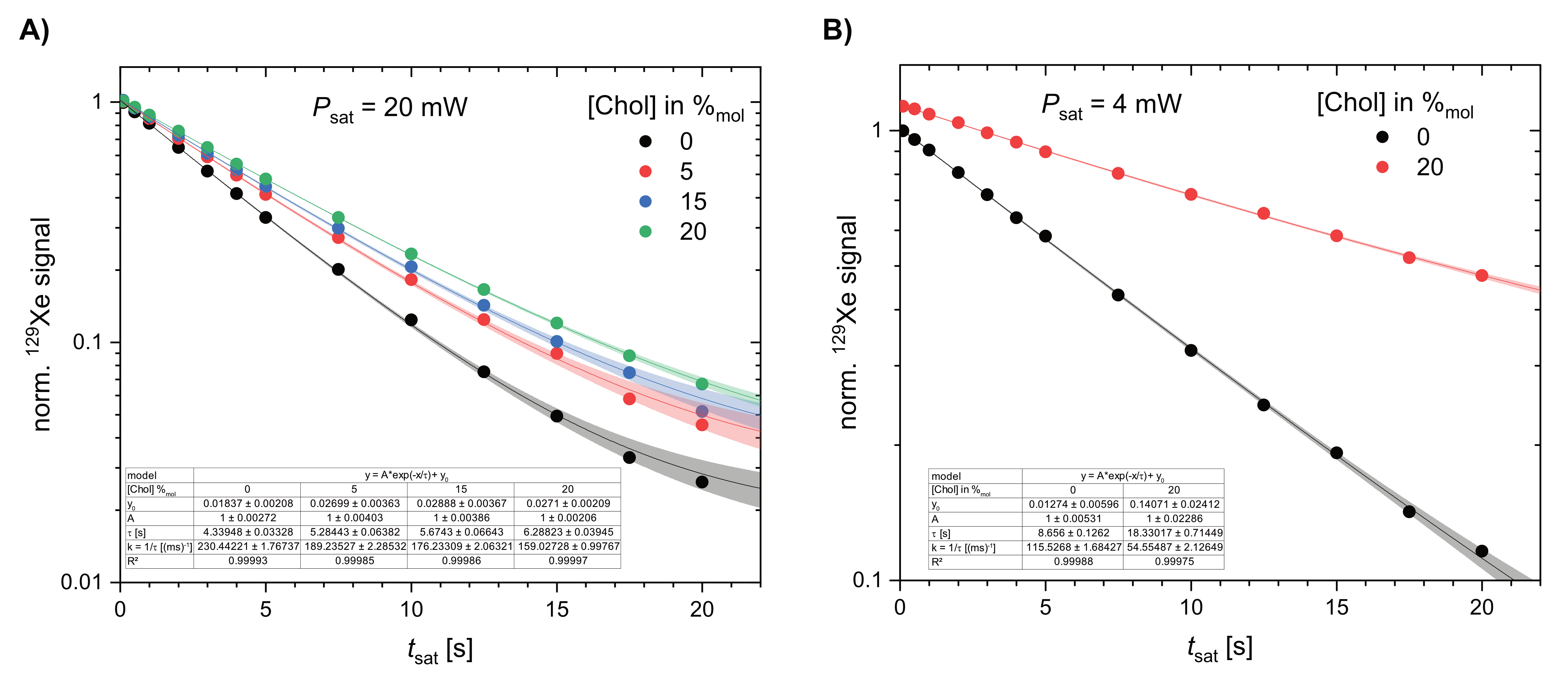
Our previously introduced lipopeptide-host conjugate was used to generate POPC-based liposomes. This lipopeptide carries cryptophane-A (CrA) as a well-studied Xe host on a Lys side chain. It thus anchors CrA inside the membrane. Cholesterol has been added at concentrations of 0, 5, 15, and 20 %mol. HyperCEST efficiency was evaluated by comparing the depolarization time constants τ to quantify the signal build-up during progressive CEST preparation with rf pulses (tsat up to 20 s at constant saturation power Psat) applied to the frequency of transiently bound Xe that is then released from the liposomes into the bulk pool. The effects were compared with POPC liposomes that carry a comparable amount of non-anchored CrA mono-acid (CrA-ma) in the presence and absence of 20 %mol cholesterol. 129Xe CEST spectroscopy was performed on a 9.4 T preclinical system with a home-made polarizer for spin exchange optical pumping and precise Xe dissolution delivery.Results and Discussion
Membrane stiffening induced by cholesterol caused a loss in observable HyperCEST efficiency for liposomes with 4 %mol CrA-lipopeptide. Fitting the depolarization time constant for the decays in Figure 2A yields results of τ = 4.34, 5.28, 5.67, 6.29 s for 0, 5, 15, 20 %mol cholesterol, respectively (Psat = 20 mW). Thus, the depolarization rate drops 1.4-fold from 230 (ms)-1 to 159 (ms)-1 upon addition of 20 %mol cholesterol. While this effect is clearly measurable, it is significantly less than for freely diffusing CrA-ma at 2 %mol concentration where the same change in cholesterol content comes with a more than 2-fold increase of the depolarization time (Figure 2B; from τ = 8.7 s to τ = 18.3 s). The depolarization rate drops from 116 (ms)-1 to 55 (ms)-1 in this case. Unfunctionalized CrA-ma was used at lower concentrations since it partially leaks out of the liposomes and causes a second slightly shifted CEST response. To minimize signal contamination from released CrA-ma, we also used a reduced saturation power (Psat = 4 mW). However, this should not impact the relative change in depolarization rate upon addition of cholesterol. We thus conclude that the membrane-anchored lipopeptide-CrA conjugate is much less susceptible to changes in membrane fluidity and retains most of its performance over a wider range of cholesterol content.This study demonstrates the superior performance of the lipopeptide approach with anchored Xe hosts. It enables better loading without host leakage and thus avoids the need to pay attention to signal contaminations from released hosts. Additional studies will investigate whether the much higher changes for CrA-ma are related to the challenge of exchanging free hosts leaving and re-entering a stiffened membrane versus the permanently membrane-anchored lipopeptide or if other effects play a role (e.g., the better host:cholesterol ratio). Overall, this study illustrates another beneficial aspect of designing HyperCEST-active liposomes with a toolbox that contains a) CrA-carrying lipopeptides in the membrane, b) targeting units on the surface, and c) various other components that tune the membrane properties for optimized circulation and HyperCEST performance.
Acknowledgements
This research was funded by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) through Koselleck Grant No. 316693477 (SCHR 995/5–1) and through support by the Dieter Morszeck Stiftung.References
[1] J. O. Jost, L. Schröder,
NMR Biomed. 2023, 36, e4714.
[2] M.
Schnurr, C. Witte, L. Schröder, Biophys. J. 2014, 106,
1301–1308.
Figures