4201
Assessing Hippocampal Energy Metabolism with Alzheimer’s Disease Progression1CSIR-Centre for Cellular and Molecular Biology, Hyderabad, India, 2Academy of Scientific and Innovative Research (AcSIR), Ghaziabad, India
Synopsis
Keywords: Alzheimer's Disease, Metabolism, 1H-[13C]-NMR, cognition, amyloid-beta plaque, glutamate, GABA, glucose
Motivation: Alzheimer’s Disease (AD) is the major form of dementia but there is no biomarker(s) and treatment available.
Goal(s): This study aimed to investigate how decline in cognitive function is manifested in neurometabolic activity with the progression of AD.
Approach: 5xFAD mice (6- and 12-month) were subjected to memory assessment using the Morris Water Maze test followed by neurometabolic analysis using 1H-[13C]-NMR spectroscopy in conjunction with an infusion of [1,6-13C2]glucose or [2-13C]acetate.
Results: The findings of the study suggest enhanced neuroinflammation, and reduced glutamatergic and GABAergic neurotransmission in 5xFAD mice.
Impact: 13C NMR based assessment of neuronal and astroglial metabolic activity may help get an insight into AD pathology, thus providing biomarker(s) for the diagnosis of AD.
Introduction
Alzheimer’s Disease (AD) is a chronic age-related neurodegenerative disorder. The common symptoms of the disease include progressive memory impairment and loss of cognitive function. AD is marked by the deposition of amyloid-beta (Aβ) plaques and neurofibrillary tangles composed of hyperphosphorylated tau1. Familial AD is the result of mutations in APP, PSEN1, and PSEN2 genes2. 5xFAD mouse model is a fast-progressing model of AD that overexpresses human APP with three AD-linked mutations and human PSEN1 with two AD-linked mutations3. In this study, we have investigated AD pathology by assessing Aβ-plaque load and neuro-metabolic activity in the hippocampus with progress of age in 5xFAD mice.Materials and Methods
Animal experiments with mice were approved by the Institutional Animal Ethics Committee of CSIR-CCMB. 5xFAD and Wild-Type (WT) mice of 6- and 12-month-olds were used in the study. The learning and memory of mice were assessed using the Morris Water Maze test4. Aβ plaque load was evaluated in formalin-fixed brain by immunohistochemical (IHC) analysis using anti-plaque antibody5. For metabolic measurement, mice were fasted for 5 h, anesthetized using urethane (1.5 mg/kg, intraperitoneal), and infused with [1,6-13C2]glucose or [2-13C]acetate through the tail vein using bolus variable infusion rates for 10 and 15 min, respectively6. The brain metabolism was arrested using a focused beam microwave irradiation (3 kW, 1.2 s) at the predefined time. The concentration and 13C labeling of metabolites were measured in 1H-[13C]-NMR spectra of hippocampal extracts recorded at 600 MHz Bruker Avance III NMR spectrometer7. The cerebral metabolic rate of glucose oxidation (CMRGlc(ox)) in glutamatergic and GABAergic neurons was calculated as described previously by Mishra et al8. The statistical significance was evaluated by Student's t-test using GraphPad Prism 8.0, and all data are presented as Mean±SD.Results
The learning of 5xFAD mice was intact at 6 months of age but became impaired by 12 months when compared to age-matched WT mice (Fig. 1A, B). The 6-month-old 5xFAD mice took a significantly longer time (p<0.01) to locate the platform (56.1±21.7 s, n=10) as compared to WT (28.1±18.2 s, n=10) mice. Similarly, at 12 months of age, 5xFAD mice took significantly more time (p=0.01) to locate the platform (68.6±28.2 s, n=7) when compared to controls (38.0±17.9s, n=8) (Fig. 1C, D). Additionally, 12-month-old 5xFAD mice spent significantly less (p=0.02) time in the platform zone (2.16±2.06 s, WT- 5.17±2.62 s) when compared to WT mice (Fig. 1E, F) during the probe test. The plaque load was significantly higher (p=0.02) in the hippocampus of 12-month-old 5xFAD mice (88.2 plaques/mm2, n=4) as compared to 6-month-old mice (48.9 plaques/mm2, n = 4) (Fig. 2A&B). The 1H-[13C]-NMR spectrum of the hippocampal extract revealed 13C labeling of amino acids (Fig. 3). The neurometabolite homeostasis was unperturbed at 6 months of age. There was a reduction in the level of glutamate (5xFAD 10.9±0.9 µmol/g; WT 11.8±0.7 µmol/g) and an increase in m-Inositol (5xFAD 8.6±1.0 µmol/g; WT 7.4±1.2 µmol/g) in the hippocampus of 12-month 5xFAD mice. The 13C label accumulated at GlnC4 (p=0.02), GluC4 (p=0.03) GABAC2 (p=0.03), and GluC3 (p=0.01) from [2-13C]acetate was increased in the hippocampus, resulting in an increased CMRAce(ox) in 12 months old 5xFAD (0.11±0.01 µmol/g/min, p=0.02) mice when compared age-matched WT controls (0.09±0.01 µmol/g/min). The labeling of GluC4 (p<0.05), GABAC2 (p<0.01), GlnC4 (p<0.05), and AspC3 (p<0.05) from [1,6-13C2]glucose in 5xFAD was reduced significantly when compared with the respective age-matched WT controls at both age groups (Table 1). Consequently, CMRGlc(ox) in Glutamatergic (5xFAD 0.27±0.06 µmol/g/min; WT 0.34±0.03 µmol/g/min, p=0.02), GABAergic neurons (0.05±0.01 vs 0.07±0.01 µmol/g/min, p<0.01), and total glucose oxidation (0.41±0.09 vs 0.52±0.05 µmol/g/min, p=0.01) was reduced significantly in 6 months old 5xFAD mice (Fig. 4). The reduction in the CMRGlc(ox) persisted till 12 months of age for CMRGlc(Glu) (0.30±0.06 vs 0.44±0.05 µmol/g/min, p<0.01) and CMRGlc(GABA) (0.08±0.01 vs 0.11±0.02 µmol/g/min, p<0.01).Discussion
The findings of the study indicated impaired learning and memory in 5xFAD mice, accompanied by an increase in Aβ-plaques in the hippocampus at 12 months. These findings are indicative of a decline in cognitive function, and accumulation of Aβ plaque, the hallmarks of AD. The increased CMRAce(ox) in 12-month-old 5xFAD mice suggests neuroinflammation in these mice. As neuronal glucose oxidation is stoichiometrically coupled with neurotransmitter cycling, the reduced CMRGlc(ox) in glutamatergic and GABAergic neurons implies a decrease in excitatory and inhibitory neurotransmission in 5xFAD mice at both ages. This was further supported by reduced labeling of GlnC4 in 5xFAD mice when compared with WT controls at both ages (Table 1).Acknowledgements
The study is supported by funding from the CSIR healthcare theme project (NCP/MLP0139). CD would like to thank DBT (DBT/2020/CCMB/1315) for Senior Research Fellowship.
References
1. Karch CM, Cruchaga C, Goate AM. Alzheimer's disease genetics: from the bench to the clinic. Neuron. 2014 Jul 2;83(1):11-26.
2. Dai MH, Zheng H, Zeng LD, Zhang Y. The genes associated with early-onset Alzheimer's disease. Oncotarget. 2017 Dec 15;9(19):15132-15143.
3. Oakley H, Cole SL, Logan S, Maus E et al. Intraneuronal beta-amyloid aggregates, neurodegeneration, and neuron loss in transgenic mice with five familial Alzheimer's disease mutations: potential factors in amyloid plaque formation. J Neurosci. 2006 Oct 4;26(40):10129-40.
4. Patel AB, Tiwari V, Veeraiah P, Saba K. Increased astroglial activity and reduced neuronal function across brain in AβPP-PS1 mouse model of Alzheimer's disease. J Cereb Blood Flow Metab. 2018 Jul;38(7):1213-1226.
5. Epstein AA, Narayanasamy P, Dash PK, High R et al. Combinatorial assessments of brain tissue metabolomics and histopathology in rodent models of human immunodeficiency virus infection. J Neuroimmune Pharmacol. 2013 Dec;8(5):1224-38.
6. Tiwari V, Ambadipudi S, Patel AB. Glutamatergic and GABAergic TCA cycle, and neurotransmitter cycling fluxes in different regions of mouse brain. J Cereb Blood Flow Metab. 2013 Oct;33(10):1523-31.
7. de Graaf RA, Brown PB, Mason GF, Rothman DL, Behar KL. Detection of [1,6-13C2]-glucose metabolism in rat brain by in vivo 1H-[13C]-NMR spectroscopy. Magn Reson Med. 2003 Jan;49(1):37-46.
8. Mishra PK, Kumar A, Behar KL, Patel AB. Subanesthetic ketamine reverses neuronal and astroglial metabolic activity deficits in a social defeat model of depression. J Neurochem. 2018 Sep;146(6):722-734.
Figures
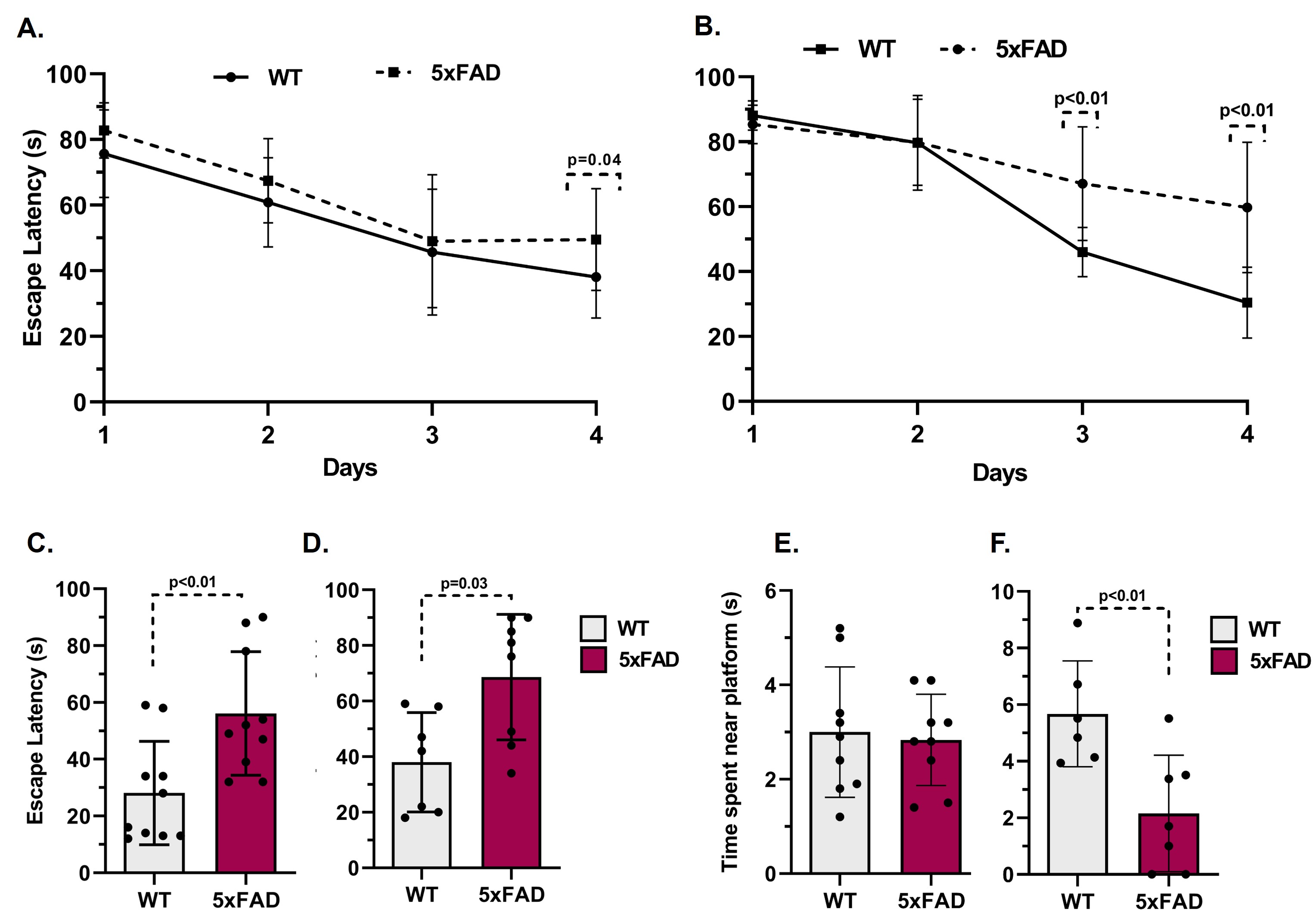
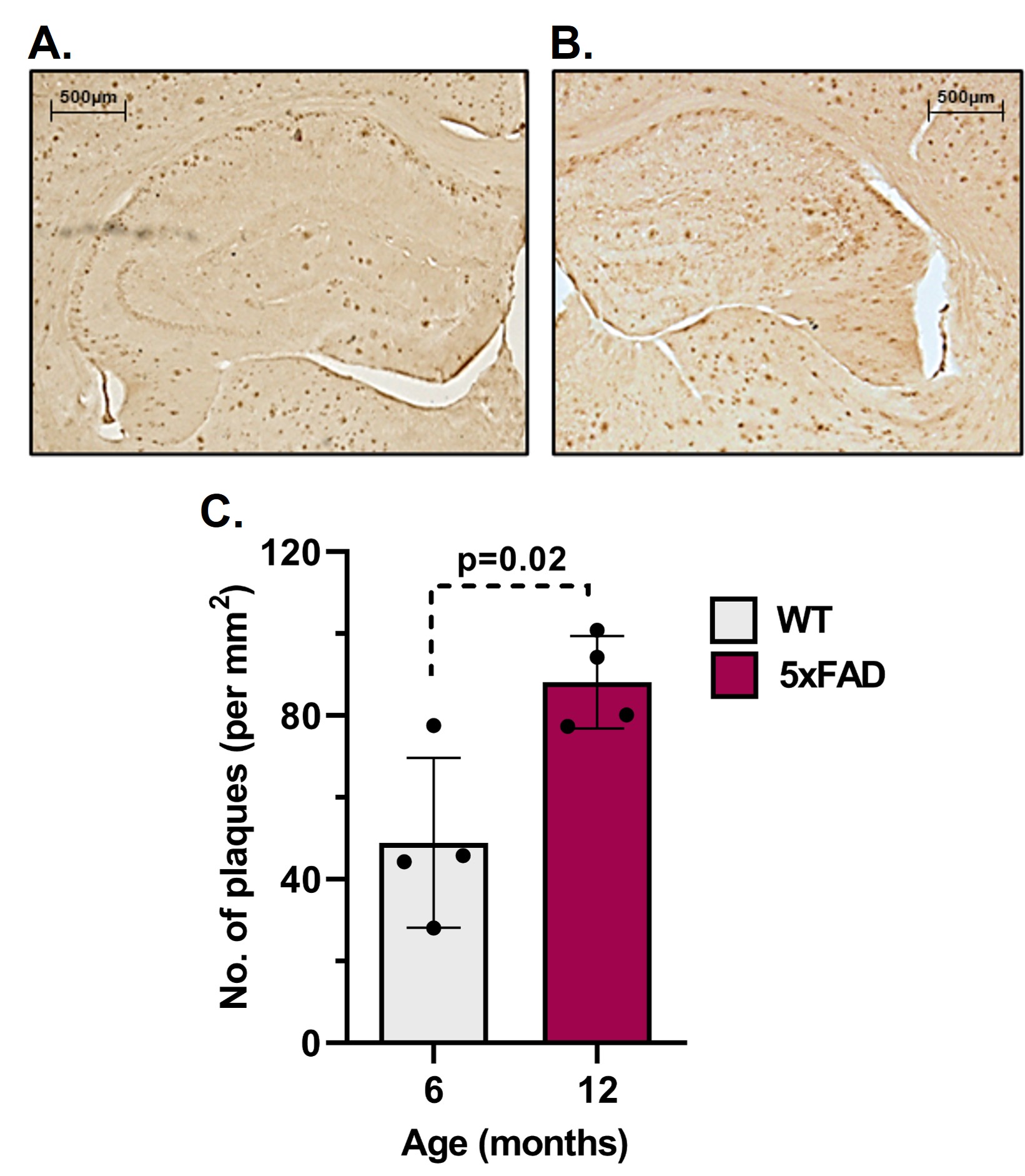
Figure 2 Plaque load in brains of A. 6 and B. 12 months old 5xFAD mice. C. Plaque count in the hippocampus of 5xFAD mice.
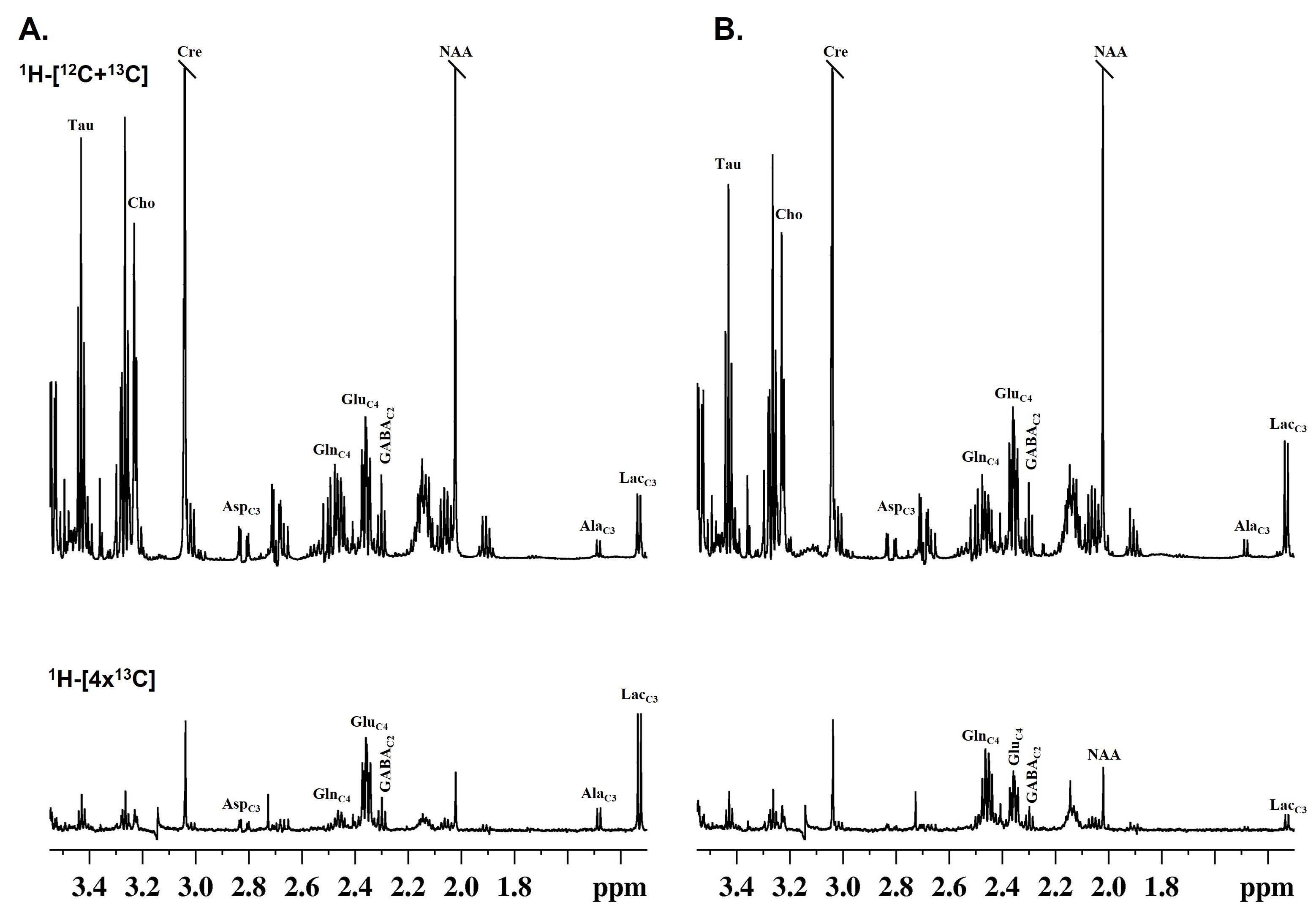
Figure 3 Representative 1H-[13C]-NMR spectra of hippocampal extract showing total and 13C labeled metabolites from A. [1,6-13C2]glucose and B. [2-13C]acetate. Mice were infused with [1,6-13C2]glucose or [2-13C]acetate followed by fixation of brain metabolism at the 10 and 15 min, respectively, using focused beam microwave irradiation. Metabolites were extracted, and 1H-[13C]-NMR spectra were recorded at 600 MHz NMR spectrometer.
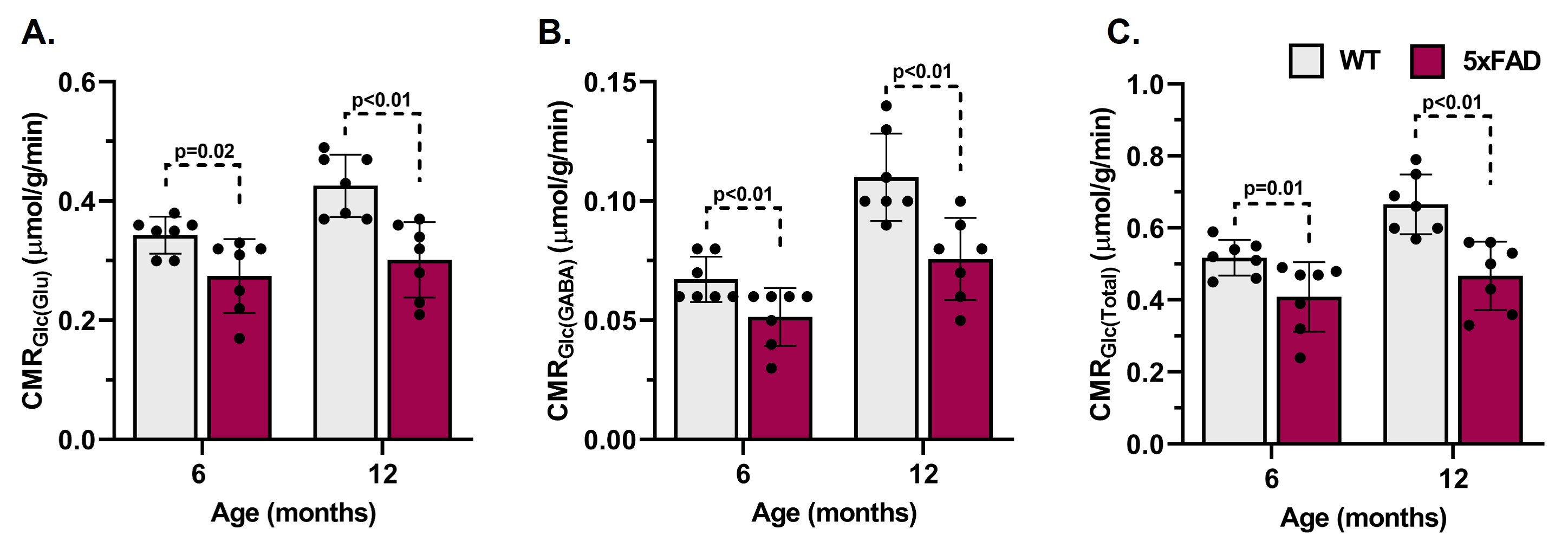
Figure 4 Cerebral metabolic rates of glucose oxidation (CMRGlc(ox)) in A. Glutamatergic neurons, B. GABAergic neurons, and C. Total in hippocampus of 6 and 12 months old 5xFAD mice. The bar graph represents the Mean±SD of the group while the circle represents measurement from individual mouse .
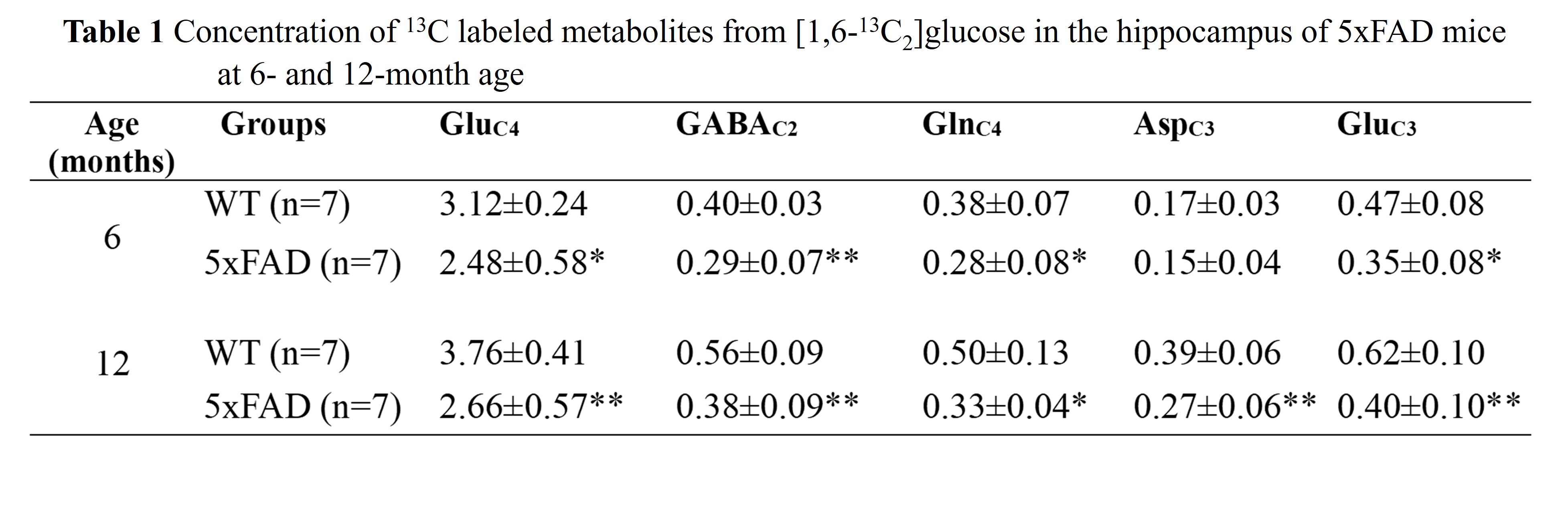
Mice were infused with [1,6-13C2]glucose followed by brain metabolism fixation at 10 min. Hippocampal metabolites were extracted, and 1H-[13C]NMR spectrum was recorded at 600 MHz NMR spectrometer. * p<0.05, ** p<0.01 compared to WT of the same age group.