4134
Optogenetic fMRI Reveals Brain-wide Functional Organization of the Zona Incerta Pathways1Laboratory of Biomedical Imaging and Signal Processing, The University of Hong Kong, Hong Kong, China, 2Department of Electrical and Electronic Engineering, The University of Hong Kong, Hong Kong, China, 3Department of Diagnostic Radiology, The University of Hong Kong, Hong Kong, China, 4School of Biomedical Sciences, The University of Hong Kong, Hong Kong, China
Synopsis
Keywords: Biology, Models, Methods, Neuroscience, fMRI Analysis, brain connectivity, preclinical
Motivation: We do not yet know where zona incerta (ZI) neural activity is distributed in central pathways despite extensive knowledge of incertal projections and functions.
Goal(s): We aim to reveal the long-range functional pathways of ZI neural activity to bridge the knowledge gap between known anatomical projections and implicated functions.
Approach: We deployed optogenetic fMRI to directly initiate neural activity at ZI and computational approaches to characterize the downstream central pathways involved.
Results: The propagation of neural activity from ZI are not restricted to monosynaptic projections within known subcortical structures, but along multi-synaptic, long-range central pathways.
Impact: Characterization of ZI functional pathways is of tremendous value to neuroscientists and clinicians for future in-depth investigations of key/previously undefined incertal regions and for designing more effective therapeutic measures such as deep brain stimulation, where ZI is a key target.
INTRODUCTION
Zona Incerta (ZI) has been implicated in diverse functional roles such as to process sensory information, regulate behaviors, convey motivational states, and participate in neural plasticity1-4. Consistent with these functions, ZI is shown to connect to nearly all major neural centers in central pathways ranging from the spinal cord, brainstem, and thalamus to the cerebral cortex5-10. Despite our extensive knowledge of the broad range of ZI anatomical projections and targets in both brain hemispheres, our understanding of where and how ZI neural activity is distributed across the central pathways to subserve these functions remains unresolved. To address this problem, it is imperative to deploy new fMRI approaches with the ability to directly initiate neural from ZI to examine the long-range incertal pathways.MATERIALS AND METHOD
Animal preparation and MRI experimental setup: 2μl rAAV5-hsyn-hChR2(H134R)-mcherry was injected into ZI in adult SD rats. After 4 weeks, an opaque optical fiber cannula (d=450μm) was implanted at the injection site (Figure 1A). All fMRI experiments were performed under 1.0% isoflurane. Blue (473nm; peak intensity = 40mW/mm2) light was presented to animals expressing ChR2 at 1 and 40 Hz (sinusoidal-shaped) in a block-design paradigm (Figure 1B).Optogenetic fMRI experiments: fMRI data was acquired on 7T Bruker scanner using GE-EPI (α=56°, TE/TR=20/1000ms). Data were preprocessed before standard GLM analysis was applied to identify significant BOLD responses.
RESULT
Frequency specific brain-wide responses driven by ZI neurons:The ZI consistently showed negative blood-oxygen-level dependent (BOLD) activations both under 1 and 40 Hz sinusoidal optogenetic stimulation. Notably, upon 1 Hz stimulation, robust and diffuse positive BOLD activations were observed both in the subcortical regions (ipsilateral SNr, MGB, dPAG, and bilateral LGN, and SC) and the cortical regions (bilateral S1BF, S2, A1, MC, V2, Cg, and PrL). When stimulation frequency was increased to 40 Hz, robust positive BOLD activations were still observed in the regions adjacent to ZI (ipsilateral SNr, MGB, and SC), but other distal brain regions showed minimal or no response (Figure 2).
Long-range Central Pathways Mediating Neural Activity Distribution Upon Zona Incerta Stimulation:
To unravel the long-range central pathways of ZI, we implemented a data-driven model (i.e., activity flow algorithm11,12; Figure 3A). Previous studies have shown that such computational approach, whereby functional network architecture identified using resting-state functional connectivity, could plausibly reflect the routes by which neural activity propagates during an overt task11,12. Here, we utilized resting-state functional connectivity of 17 age-matched rats (i.e., with optical fiber in ZI) as the connectivity weights in the model (Figure 3B) to predict the activations caused by ZI stimulation. Overall, the model was able to predict the activations. However, we observed aberrant predictions at several subcortical and cortical regions, indicating pathways downstream from ZI that were significantly engaged upon optogenetic stimulation (Figure 3C). Subsequently, through further K-Means cluster analysis, with the clustering criterion set as the mean absolute error (MAE) between actual and predicted activations, we discovered two clusters (orange and blue) of brain regions (A1L/R, S1ULpL, S2R, M1R, CgL, PrLL/R, ICR, MGBR, SCL/R, dPAG, GPVPR, CPuL/R, InsR, OFCL, ECR, and dHPR) with high MAE under 1 Hz stimulation, while only clusters in the subcortical regions (Sth-SNr-TgL/R, SCR and MGBR, Figure 3D) showed high MAE under 40 Hz stimulation.
LFPs recordings in ZI and dPAG confirm neuronal activity underlying BOLD activations:
Guided by our fMRI findings, we conducted LFPs recordings using the same stimulation paradigm to investigate the underlying neural activity induced by optogenetic stimulation in ZI. Through time-frequency/spectrogram analysis13 of the LFPs responses recorded in ZI and dorsal periaqueductal gray (dPAG), we found that evoked neural activity in ZI followed the sinusoidal optogenetic stimulus (Figure 4A) and gamma oscillations were induced in dPAG upon 1 Hz stimulation in ZI (Figure 4B).
DISSCUSION AND CONCLUSION
Although ZI has minimal direct projections to the cortex6, our results showed that it can engage several brain regions that in turn project to the cortex, such as SNr, LGN, and MGB16, indicating that ZI can modulate cortical activity through indirect projections. Notably, through our modeling approach, we found strengthened downstream ZI pathways that were associated with memory functions, auditory information processing, and modulation of fear-related behaviors. A subset of these distinct pathways within the sensorimotor network engaged by ZI’s activation could be mediating the sound-induced innate flight and freezing behaviors in a recent work on ZI functions2. In conclusion, our results demonstrate that the neural activity propagation from ZI neurons are not restricted to monosynaptic projections from the ZI to known subcortical structures, but along multi-synaptic, long-range central pathways, specifically at low frequencies.Acknowledgements
This work was supported in part by Hong Kong Research Grant Council (HKU17112120, HKU17127121, HKU17127022 and HKU17127523 to E.X.W. and HKU17104020, HKU17127021, HKU17127723 to A.T.L.L.), Lam Woo Foundation, and Guangdong Key Technologies for AD Diagnostic and Treatment of Brain (2018B030336001) to E.X.W.References
1. Wang X, Chou XL, Zhang LI, Tao HW. Zona Incerta: An Integrative Node for Global Behavioral Modulation. Trends Neurosci. 2020. 43(2):82-87.
2. Chou XL, Wang X, Zhang ZG, Shen L, Zingg B, Huang J, Zhong W, Mesik L, Zhang LI, Tao HW. Inhibitory gain modulation of defense behaviors by zona incerta. Nat Commun. 2018. 9(1):1151.
3. Zhao ZD, Chen Z, Xiang X, Hu M, Xie H, Jia X, Cai F, Cui Y, Chen Z, Qian L, Liu J, Shang C, Yang Y, Ni X, Sun W, Hu J, Cao P, Li H, Shen WL. Zona incerta GABAergic neurons integrate prey-related sensory signals and induce an appetitive drive to promote hunting. Nat Neurosci. 2019. 22(6):921-932.
4. Liu K, Kim J, Kim DW, Zhang YS, Bao H, Denaxa M, Lim SA, Kim E, Liu C, Wickersham IR, Pachnis V, Hattar S, Song J, Brown SP, Blackshaw S. Lhx6-positive GABA-releasing neurons of the zona incerta promote sleep. Nature. 2017. 548(7669):582-587.
5. Watson C, Lind CR, Thomas MG. The anatomy of the caudal zona incerta in rodents and primates. J Anat. 2014. 224(2):95-107.
6. Yang Y, Jiang T, Jia X, Yuan J, Li X, Gong H. Whole-Brain Connectome of GABAergic Neurons in the Mouse Zona Incerta. Neurosci Bull. 38(11):1315-1329.
7. Gan Z, Gangadharan V, Liu S, Körber C, Tan LL, Li H, Oswald MJ, Kang J, Martin-Cortecero J, Männich D, Groh A, Kuner T, Wieland S, Kuner R. Layer-specific pain relief pathways originating from primary motor cortex. Science. 2022. 378(6626):1336-1343.
8. Cavdar S, Onat F, Cakmak YO, Saka E, Yananli HR, Aker R. Connections of the zona incerta to the reticular nucleus of the thalamus in the rat. J Anat. 2006 Aug;209(2):251-8.
9. Chen J, Kriegstein AR. A GABAergic projection from the zona incerta to cortex promotes cortical neuron development. Science. 2015 Oct 30;350(6260):554-8.
10. Lin CS, Nicolelis MA, Schneider JS, Chapin JK. A major direct GABAergic pathway from zona incerta to neocortex. Science. 1990. 248(4962):1553-6.
11. Cole MW, Ito T, Bassett DS, Schultz DH. Activity flow over resting-state networks shapes cognitive task activations. Nat Neurosci. 2016. 19(12):1718-1726.
12. Hearne LJ, Mill RD, Keane BP, Repovš G, Anticevic A, Cole MW. Activity flow underlying abnormalities in brain activations and cognition in schizophrenia. Sci Adv. 2021. 7(29):eabf2513.
13. Arts, L.P.A., van den Broek, E.L. The fast continuous wavelet transformation (fCWT) for real-time, high-quality, noise-resistant time–frequency analysis. Nat Comput Sci. 2022. 2: 47–58.
14. Canolty RT, Knight RT. The functional role of cross-frequency coupling. Trends Cogn Sci. 2010. 14(11):506-15.
15. Oh SW, Harris JA, Ng L, Winslow B, Cain N, Mihalas S, Wang Q, Lau C, Kuan L, Henry AM, Mortrud MT, Ouellette B, Nguyen TN, Sorensen SA, Slaughterbeck CR, Wakeman W, Li Y, Feng D, Ho A, Nicholas E, Hirokawa KE, Bohn P, Joines KM, Peng H, Hawrylycz MJ, Phillips JW, Hohmann JG, Wohnoutka P, Gerfen CR, Koch C, Bernard A, Dang C, Jones AR, Zeng H. A mesoscale connectome of the mouse brain. Nature. 2014. 508(7495):207-14.
16. Loughlin SE, Fallon JH. Substantia nigra and ventral tegmental area projections to cortex: topography and collateralization. Neuroscience. 1984. 11(2):425-35.
Figures
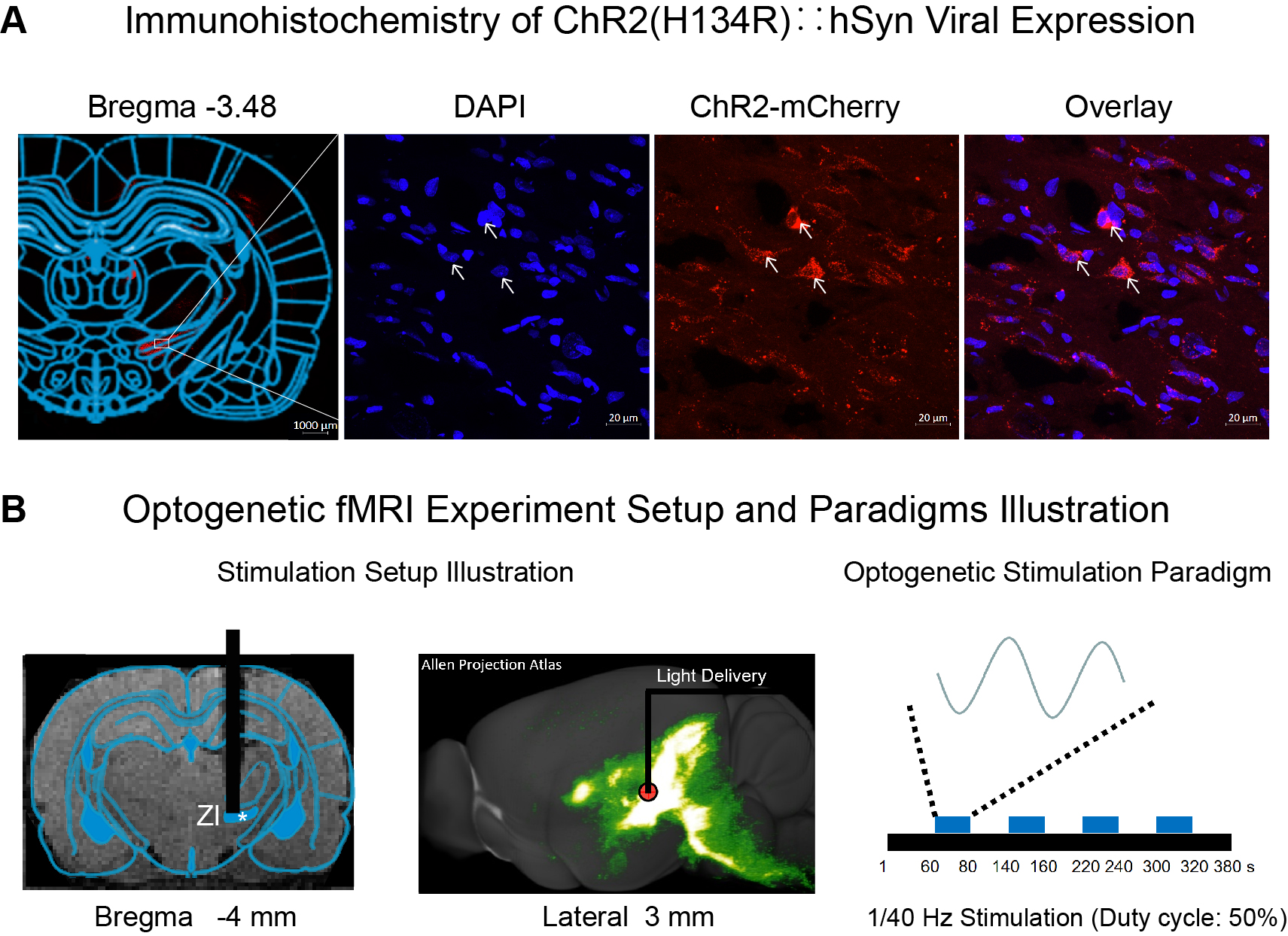
Fig.1 Histological characterization of ChR2::hSyn viral expression in zona incerta (ZI) neurons and optogenetic fMRI stimulation setup.
(A) Confocal images of ChR2-mCherry expression in ZI. Overlay of images costained for the nuclear marker DAPI and mCherry revealed colocalization of ChR2 in the cell body of ZI neurons (indicated by white arrows). (B) Illustration of stimulation site in ZI (left). The 3D projection schematic diagram is from the public database of the Allen Institute (middle)15. Optogenetic fMRI stimulation paradigm (right).
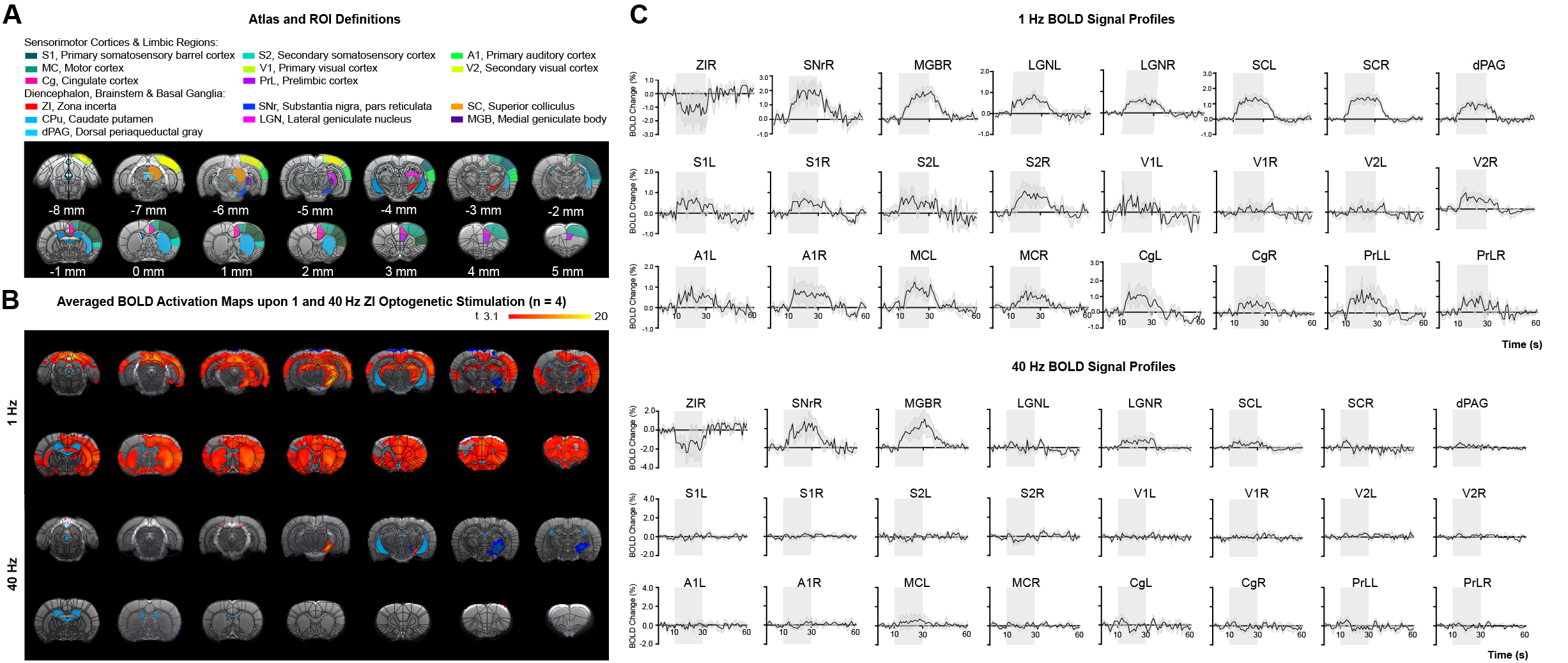
Fig.2 Low-frequency (1 Hz), not high-frequency (40 Hz), optogenetic excitation of ZI neurons evokes brain-wide positive BOLD activations in remote and bilateral sensory and high-order cortices.
(A) Illustration of Paxinos atlas-based ROI definitions. (B) Averaged BOLD activation maps at 1 and 40 Hz optogenetic stimulation (n = 4). (C) BOLD signal profiles extracted from ROIs defined in A (error bar indicates ± SEM).
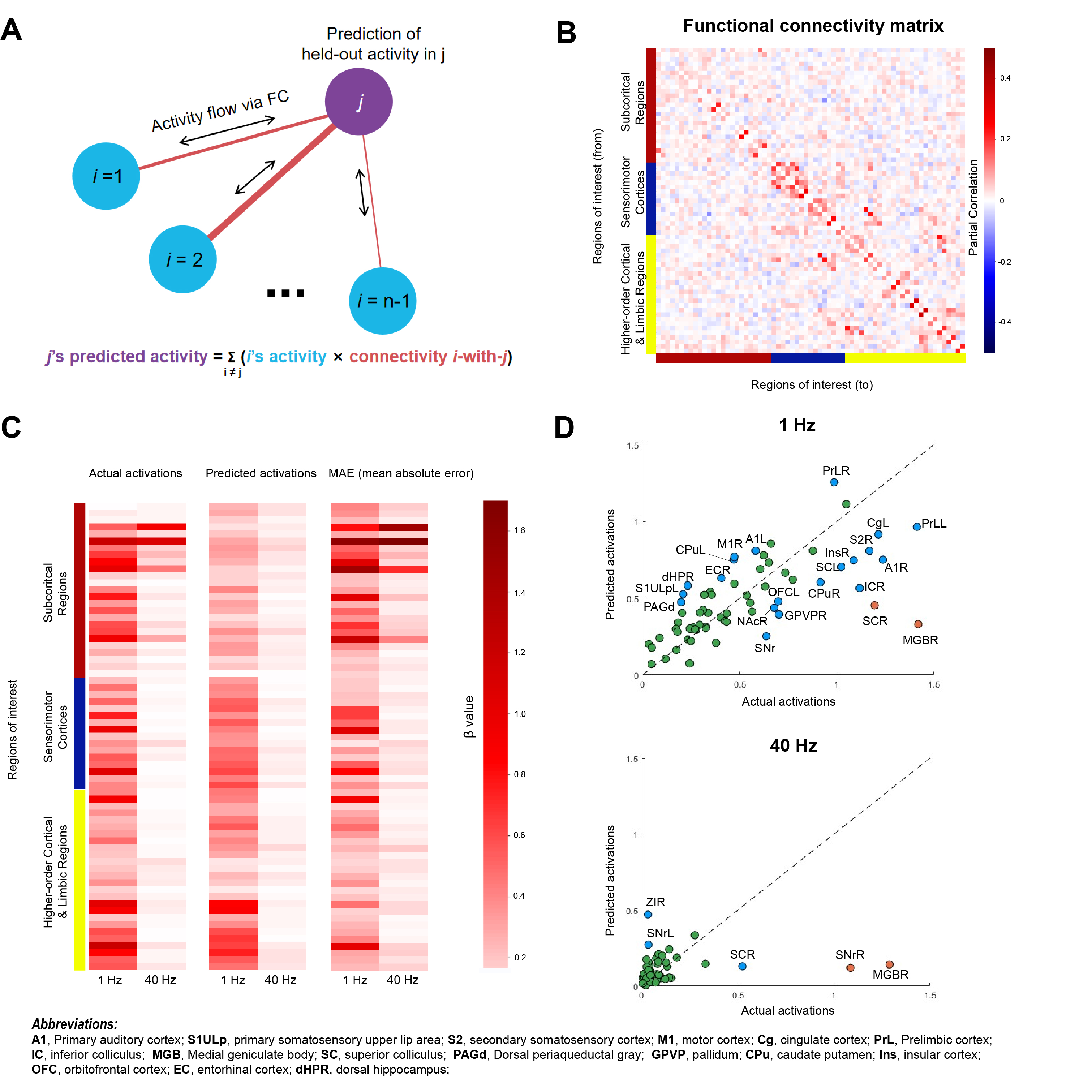
Fig. 3 Predicting optogenetically evoked activity with activity flow algorithm.
(A) The activity flow algorithm11-12. The task-evoked activation of brain region (j) can be predicted by summing the activity of all other brain regions (i) weighted by their connectivity with j. (B) Partial correlation matrix computed based on resting-state fMRI data (n = 17). (C) Group averaged actual, predicted activations, and MAE for 1 Hz and 40 Hz optogenetic fMRI data. (D) K-Means cluster analysis with three distinct clusters identified. The dashed line in the plot represents the line y=x.
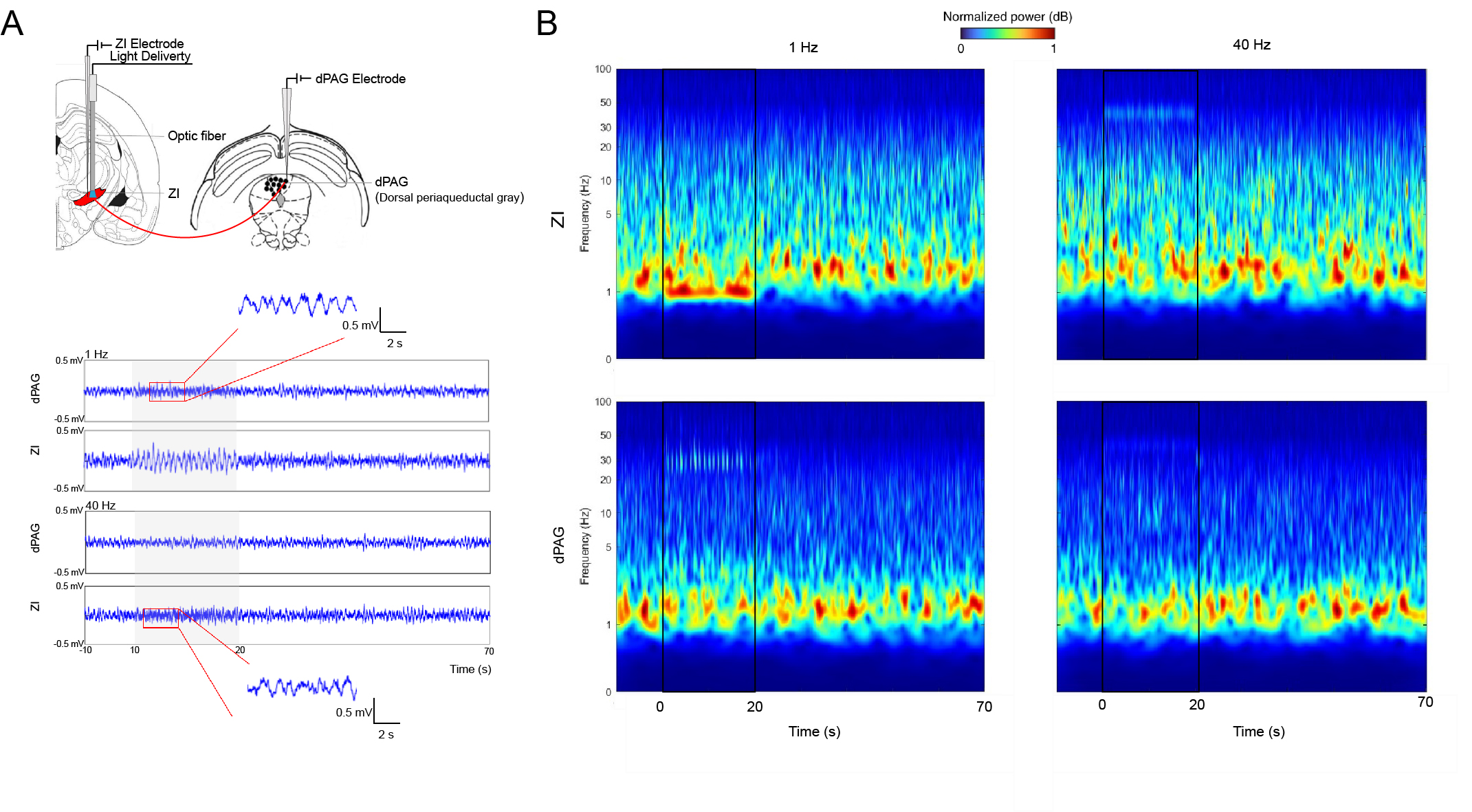
Fig. 4 LFPs recordings in ZI and dPAG confirm neuronal activity underlying BOLD activation.
(A) Illustration of the location of recording electrodes in ZI and dPAG (Top). Single-channel tungsten microelectrodes (1 MΩ and 10-μm tip diameter) were placed in ZI and dPAG to record local field potentials (LFPs) in animals using the same protocol as in MRI experiments. Averaged LFPs from a representative animal during ZI optogenetic stimulation (n=3, bottom). (B) Time-frequency analysis of LFPs recorded in ZI and dPAG.