4128
The hippocampus CA1 microcircuit is ready to join the taskforce of region-specific mean field models for a comprehensive virtual brain simulation1Department of Brain and Behavioral Sciences, Università di Pavia, Pavia, Italy, 2Paris-Saclay University, CNRS, Paris-Saclay Institute of Neuroscience,Gif-sur-Yvette, Paris, France, 3Digital Neuroscience Centre, IRCCS Mondino Foundation, Pavia, Italy, 4NMR Research Unit, Queen Square MS Centre, Department of Neuroinflammation, UCL Queen Square Institute of Neurology, Faculty of Brain Sciences, University College London, London, United Kingdom, 5Institute of Biophysics, National Research Council, Palermo, Italy
Synopsis
Keywords: Biology, Models, Methods, Neuroscience, Modelling, new device, hippocampus
Motivation: Virtual brains are proving quite successful for BOLD signal simulation but they still lack a specific mathematical description of individual brain regions.
Goal(s): Our goal was to develop a specific mean field model (MF) of the hippocampus CA1 microcircuit.
Approach: A bottom-up formalism recently developed for the cerebral and cerebellar cortex was adopted. It is based on a transfer function, which remaps neuronal microscale features from cellular recordings to the mean field mesoscale domain.
Results: The hippocampus CA1 MF reproduced the neuronal activity of the microcircuit and captured learning mechanisms.
Impact: The mean field of the hippocampus CA1 microcircuit enriches the growing collection of region-specific models. Its forthcoming integration into virtual brains together with other region-specific models has the potential to achieve a comprehensive and personalised BOLD signal dynamics simulation.
Introduction
Region-specific models are the fundamental bricks needed to generate effective virtual brain models and digital twins1,2. However, at present virtual brains still use generic “top-down” neural mass models, which impose a strong limitation on their precision. Recently, “bottom-up” mean field models (MFs) have been proposed for the cerebral and cerebellar cortices. They have been constructed using detailed cortical microcircuit simulations validated on preclinical cellular data, and therefore account for microstructural and electrophysiological properties of corresponding brain regions3. Here we exploited the flexibility of this bottom-up MF formalism to develop a MF model of the hippocampal CA1 microcircuit (CA1 MF). The hippocampus plays a key role in brain functions such as learning, memory consolidation and spatial navigation4–6. CA1 MF predictions were tested against the corresponding microscale simulations (i.e., those obtained with the CA1 spiking neural network (SNN)) to generate rhythmic activity and synaptic transmission patterns.Methods
The CA1 MF pipeline is reported in Figure 1. Model architecture (Figure 2) relied on a previously validate microcircuit model7. The CA1 MF circuit was assumed to be constituted by two populations, i.e., “regular spiking” pyramidal excitatory neurons (RS)8 and “fast spiking” inhibitory interneurons (FS)9. The external input, coming from the hippocampal CA3 region, targets both FS and RS cells.The CA1 MF was developed following an already well assessed approach based on a Transfer Function (F) formalism that transfers neuronal microscale parameters, estimated experimentally, into MF equations10. A specific F was computed for RS and FS (i.e., FRS and FFS) incorporating connection probabilities, synaptic decay times and quantal conductances (K, τ, Q) extracted from validated SNN, and providing as output the average population activity. The procedure yielded a numerical table containing the firing-frequency of the neuronal population (numerical F)10. The analytical form of F was then obtained by a fitting procedure on the numerical F (Figure 3). The MF time constant (T) was set at 5 ms10.
CA1 MF predictions were compared to the SNN outputs, setting the external input so as to explore different physiological activity regimes observed in CA111,12. CA1 MF predictive validity of complex synaptic mechanisms, such as learning condition, was investigated by tuning the weight of synaptic connection between RS and the external input, to emulate different conditions induced by Long-Term Depression and Potentiation (LTD/LTP)13,14.
Results
CA1 MF captured the dynamic activity of the SNN, both for sinusoidal inputs in the theta and gamma bands and for gaussian-shaped inputs emulating sharp waves (Figure 4).Simulations exploring RS learning showed a different CA1 MF activity according to the variation in synaptic weights between RS and external inputs (Figure 5A). RS activity showed a nonlinear trend, increasing from LTD to LTP conditions (Figure 5B).
Discussion
The CA1 MF was developed adopting the same transfer function formalism previously used for MFs of the cerebral and cerebellar cortices3,15. CA1 MF can correctly reproduce activity patterns of the corresponding SNN including theta and gamma oscillations, and sharp-waves.Synaptic plasticity in the hippocampus, especially within the synapses of RS cells, has been extensively studied, revealing its pivotal role in learning and memory processes. The nonlinear trend observed in RS activity, from LTD to LTP, shows that the effects of plasticity can be captured by MF predictions16. However, synaptic learning rules are not explicitly included in the MF formalism, but they could be integrated in the SNN. Therefore, further exploration of LTD/LTP conditions could be achieved by including learning rules in the SNN and comparing its output with MF predictions. The process of parameter tuning not only demonstrates the model adaptability but also paves the way for further manipulation to remap physiological and potentially pathological features onto the CA1 MF. In future work, efforts will focus on identifying and extracting biophysically meaningful features from subject-specific data such as diffusion-weighted imaging, enabling the customization of the CA1 MF to individual characteristics. This combination of functional simulations with (micro)structural information promises to advance our understanding of hippocampal function in subject-specific context.
Furthermore, the CA1 MF was developed to be integrated in brain dynamic simulators, enabling us to bridge the gap between micro and macroscale dynamics exploiting its across-scales nature. Its connection with other cortical and subcortical MFs could bring us closer to the realization of a comprehensive digital twin of the brain1,17.
Conclusion
The CA1 MF is ready to join the “taskforce” of transfer function-based MF models, and their integration into a virtual brain simulator will make BOLD simulations directly linked with region-specific microscale physiological properties, significantly advancing our understanding of brain function and dysfunction.Acknowledgements
This research has received funding from the European Union’s Horizon 2020 Framework Program for Research and Innovation under the Specific Grant Agreement No. 945539 (Human Brain Project SGA3) to ED, AD, CGWK, and FP. CGWK received funding from BRC (#BRC704/CAP/CGW), MRC (#MR/S026088/1), Ataxia UK, Rosetree trust (#PGL22/100041 and #PGL21/10079). CGWK is a shareholder in Queen Square Analytics Ltd. This research has also received funding from Centro Fermi project “Local Neuronal Microcircuits” to ED. Special acknowledgement to EBRAINS and FENIX for informatic support and infrastructure. RL have been supported by Human Brain Project SGA3. This work was also supported by #NEXTGENERATIONEU (NGEU) and funded by the Ministry of University and Research (MUR), National Recovery and Resilience Plan (NRRP), project MNESYS (PE0000006) – A Multiscale integrated approach to the study of the nervous system in health and disease (DN. 1553 11.10.2022) to ED, CGWK and CC; and Project EBRAINS-Italy (IR00011) - (M4C2 Line 3.1 of the PNRR, Action 3.1.1 - CUP B51E22000150006) to ED and CC.
References
[1] D’Angelo E, Jirsa V. Neurosciences The quest for multiscale brain modeling. Trends Neurosci. 2022;xx(xx):1-14. doi:10.1016/j.tins.2022.06.007
[2] Sanz leon P, Knock SA, Woodman MM, et al. The virtual brain: A simulator of primate brain network dynamics. Front Neuroinform. 2013;7(MAY). doi:10.3389/fninf.2013.00010
[3] Lorenzi RM, Geminiani A, Zerlaut Y, et al. A multi-layer mean-field model of the cerebellum embedding microstructure and population-specific dynamics. PLoS Comput Biol. 2023:1-30. doi:10.1371/journal.pcbi.1011434
[4] Buzsáki G. Two-stage model of memory trace formation: a role for “noisy” brain states. Neuroscience. 1989;31(3):551-570. doi:10.1016/0306-4522(89)90423-5
[5] RANCK JB. The Hippocampus as a Cognitive Map. Vol 136.; 1979. doi:10.1176/ajp.136.10.1353
[6] Bird CM, Burgess N. The hippocampus and memory: insights from spatial processing. Nat Rev Neurosci. 2008;9(3):182-194. doi:10.1038/nrn2335
[7] Marasco A, Spera E, De Falco V, et al. An Adaptive Generalized Leaky Integrate-and-Fire Model for Hippocampal CA1 Pyramidal Neurons and Interneurons. Bull Math Biol. 2023;85(11):109. doi:10.1007/s11538-023-01206-8
[8] Migliore R, Lupascu CA, Bologna LL, et al. The physiological variability of channel density in hippocampal CA1 pyramidal cells and interneurons explored using a unified data-driven modeling workflow. PLoS Comput Biol. 2018;14(9):1-25. doi:10.1371/journal.pcbi.1006423
[9] Sik A, Penttonen M, Ylinen A, Buzsáki G. Hippocampal CA1 interneurons: an in vivo intracellular labeling study. J Neurosci Off J Soc Neurosci. 1995;15(10):6651-6665. doi:10.1523/JNEUROSCI.15-10-06651.1995
[10] Zerlaut Y, Chemla S, Chavane F, Destexhe A. Modeling mesoscopic cortical dynamics using a mean-field model of conductance-based networks of adaptive exponential integrate-and-fire neurons. J Comput Neurosci. 2018;44(1):45-61. doi:10.1007/s10827-017-0668-2
[11] Skinner FK, Rich S, Lunyov AR, Lefebvre J, Chatzikalymniou AP. A Hypothesis for Theta Rhythm Frequency Control in CA1 Microcircuits. Front Neural Circuits. 2021;15(April):1-11. doi:10.3389/fncir.2021.643360
[12] Mysin I. A Model of the CA1 Field Rhythms. eNeuro. 2021;8(6). doi:10.1523/ENEURO.0192-21.2021
[13] Malenka RC. Synaptic plasticity in the hippocampus: LTP and LTD. Cell. 1994;78(4):535-538. doi:10.1016/0092-8674(94)90517-7
[14] Gonçalves-Ribeiro J, Pina CC, Sebastião AM, Vaz SH. Glutamate Transporters in Hippocampal LTD/LTP: Not Just Prevention of Excitotoxicity. Front Cell Neurosci. 2019;13(August):1-7. doi:10.3389/fncel.2019.00357
[15] Di Volo M, Romagnoni A, Capone C, Destexhe A. Biologically Realistic Mena-Field Models of Conductance-Based Networks of Spiking Neurons with Adaptation. Neural Comput. 2019;31:653-680. doi:10.1162/neco_a_01173
[16] O’Reilly RC, Rudy JW. Conjunctive representations in learning and memory: Principles of cortical and hippocampal function. Psychol Rev. 2001;108(2):311-345. doi:10.1037/0033-295X.108.2.311
[17] Deco G, Tononi G, Boly M, Kringelbach ML. Rethinking segregation and integration: contributions of whole-brain modelling. Nat Rev Neurosci. 2015;16(7):430-439. doi:10.1038/nrn3963
Figures
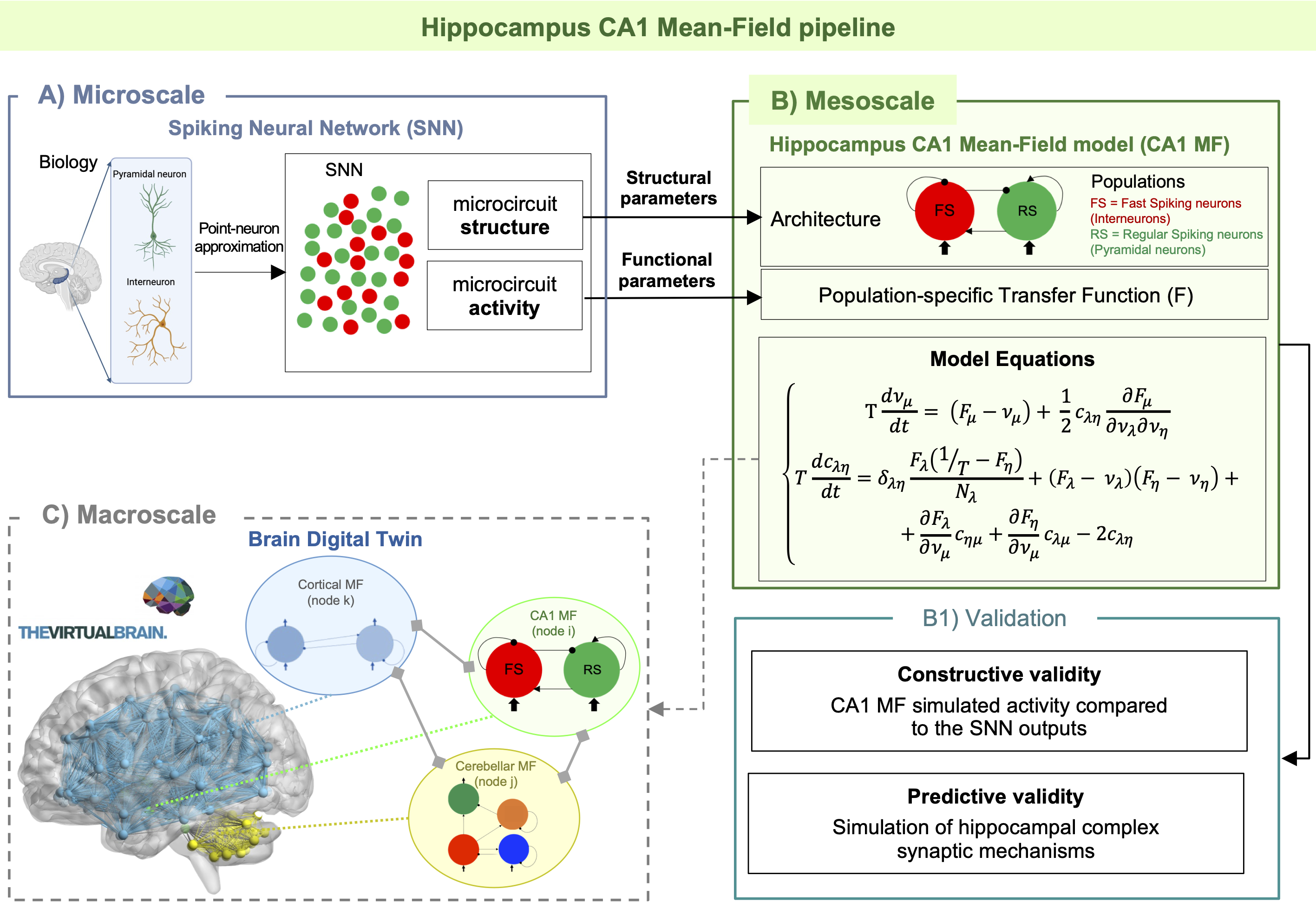
Figure 1) CA1 Mean Field (MF) Pipeline. A) Scheme of how a MF model is constructed using structural and functional parameters from spiking neural networks. B) Model equations reproducing mean activity (ν [Hz]) and variance (c [Hz]) for each population with their presynaptic connections (μ, η, λ = [FS, RS]). δ = Dirac function. B1) Model validation with observed output and tested to predict hippocampal learning. C) Next step towards a brain digital twin, with the integration of the MF into virtual brains simulators, connected to other models.
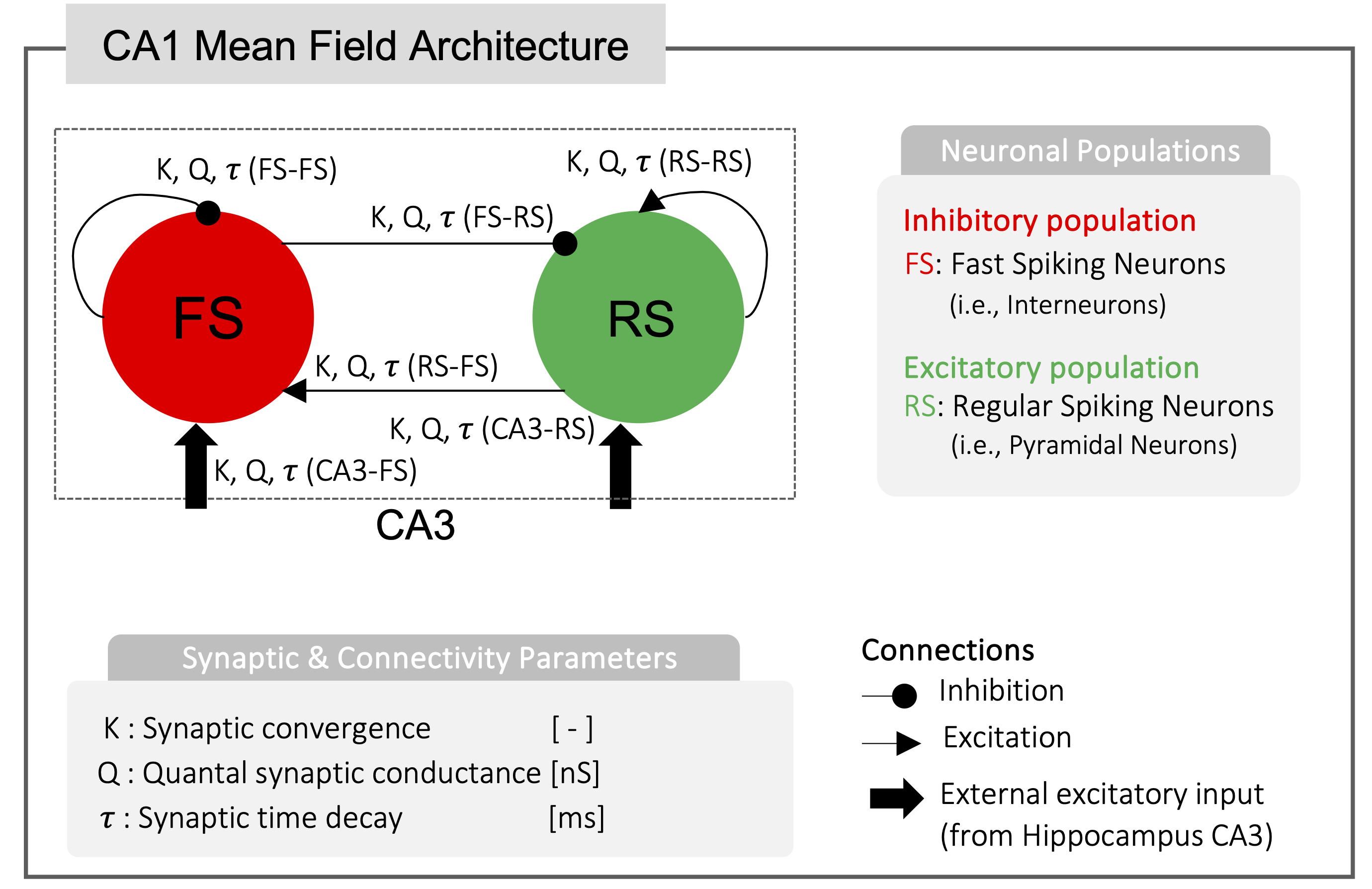
Figure 2) CA1 Mean Field (MF) Architecture. One excitatory and one inhibitory population are included: excitatory regular spiking pyramidal neurons (RS, green) and fast spiking inhibitory interneurons (FS, red). The CA1 MF receives an external input from the Hippocampus CA3 microcircuit. Architecture design and synapses-specific connectivity parameters (K, Q, and t) were extracted form previously validated microcircuit model and spiking neural network simulations, accurately reproducing cellular behavior in preclinical setting7.
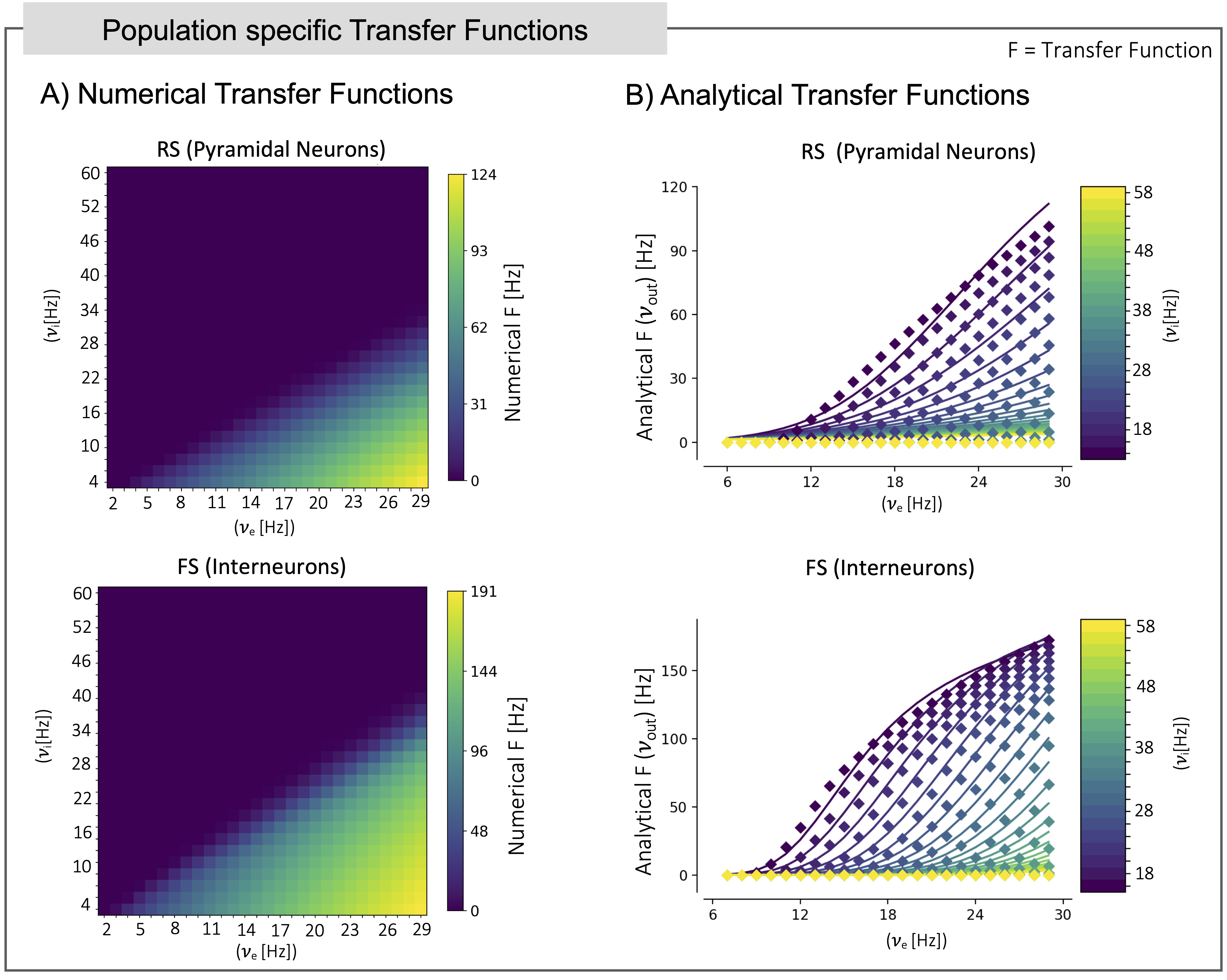
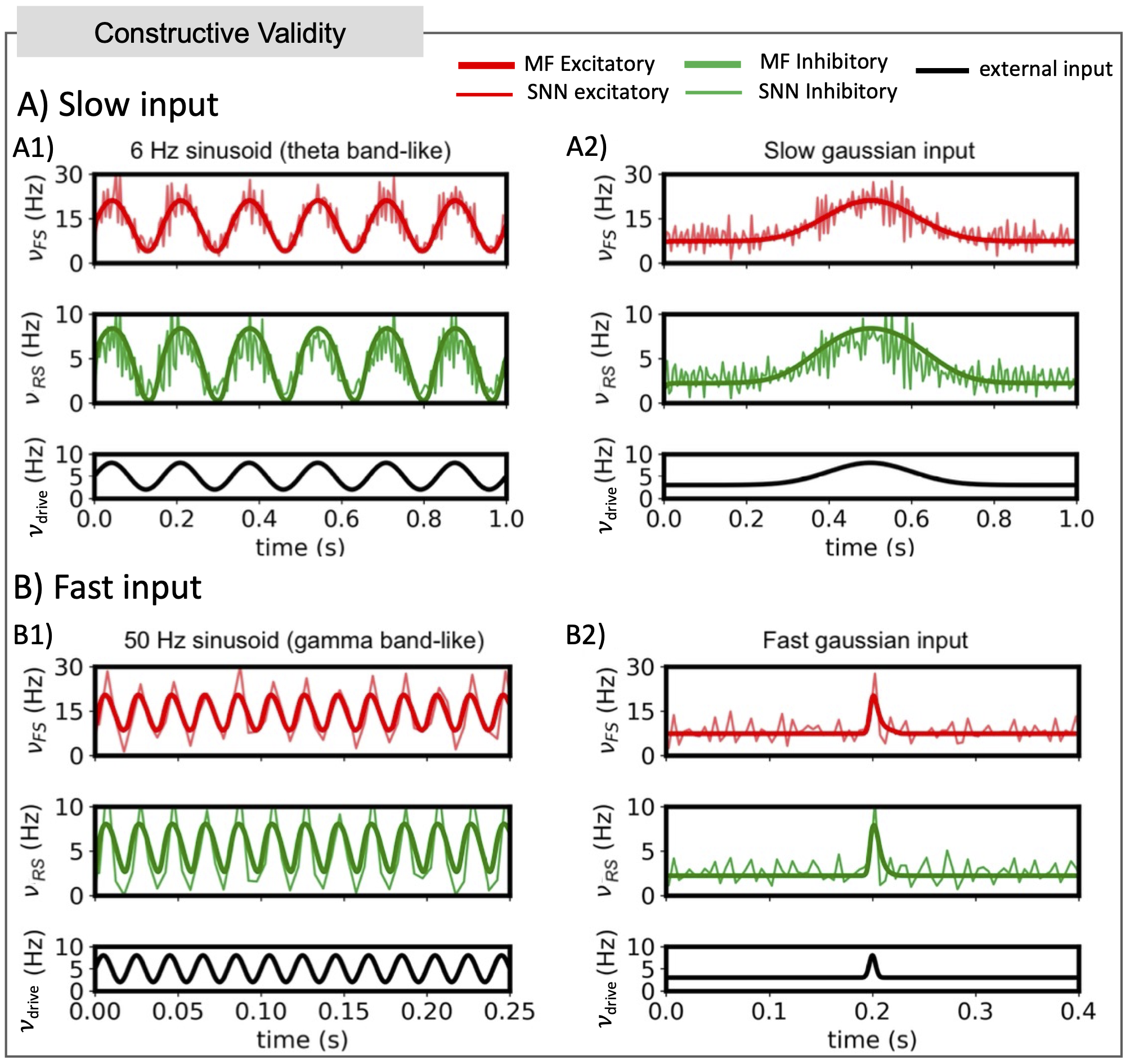
Figure 4) Constructive validity: Comparison between mean field (MF) prediction and spiking neural network (SNN). MF results (bold solid lines) are superimposed to the activity from the spiking SNN simulations of the hippocampus (light solid lines). Response of the system is reported for slow (A) and fast input (B). The MF captures the response of the SNN in large frequency-ranges, both for sinusoidal and gaussian-shaped inputs, that are relevant to simulate different activity patterns observed in the hippocampus.
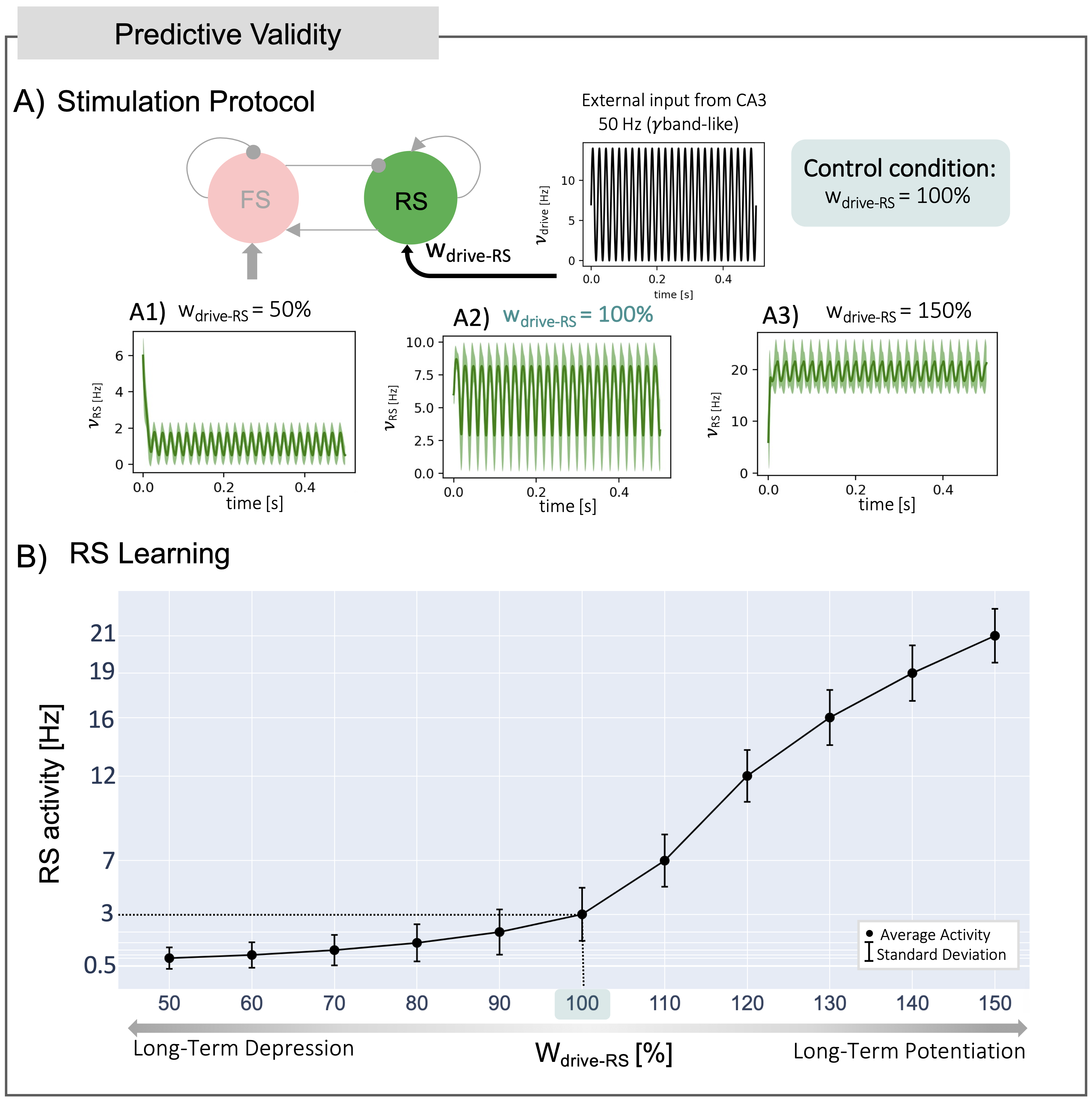
Figure 5) Predictive Validity: Long-Term Depression/Potentiation (LTD/LTP) conditions simulated by tuning the synaptic weights between the external input (drive) and RS cells (wdrive-RS). A) Simulation Protocol: wdrive-RS defined from 50% (LTD) to 150% (LTP) of the control condition (i.e., wdrive-RS = 100%) (Panels A1-3). For each wdrive-RS, a simulation of 500 ms with 50 Hz sinusoidal input was run. B) RS Learning: RS activity at steady state has a nonlinear trend for each level of wdrive-RS from LTD to LTP, capturing the synaptic complexity at the basis of learning.