4126
Brain metabolism in mice after RhoGTPases pharmacological modulation: a MRI and MRS study1MRI Unit, Core Facilities, Istituto Superiore di Sanita', Rome, Italy, 2Department of Basic and Applied Sciences for Engineering, Sapienza University of Rome, Rome, Italy, 3Department of Cardiovascular, Endocrine-Metabolic and Ageing-Associated Diseases, Istituto Superiore di Sanita', Rome, Italy, 4Department of Physiology and Pharmacology “V. Erspamer, Sapienza University of Rome, Rome, Italy, 5Center Behavioral Sciences and Mental Health, Istituto Superiore di Sanita', Rome, Italy
Synopsis
Keywords: Preclinical Image Analysis, Spectroscopy, animals, aging, spectroscopy, treatment, Rho GTPases
Motivation: RhoGTPases are involved in the pathogenesis of neurodegenerative and neurodevelopmental diseases, playing a key role in synaptic plasticity.
Goal(s): The aim is to evaluate the involvement of RhoGTPases on ageing processes, assessing effects of RhoGTPases modulation on behavior and brain metabolism (by MRS) on a healthy aging mouse model.
Approach: We pharmacologically modulated RhoGTPases pathways by administering fasudil or CNF1 in mice. Animals are subjected to behavioral tests and to MRI/MRS experiments in frontal cortex, hippocampus and cerebellum.
Results: Young mice showed apparent greater anxiolytic effect respect to old mice. We observed also age- differences in the metabolism in response to both treatments.
Impact: Dysregulation of the RhoGTPase pathway leads to: oxidative stress, inflammation, alterations in energy metabolism and reduced synaptic transmission. RhoGTPase pathway is involved in the pathogenesis of neurodegenerative diseases; RhoGTPases are here proposed as an innovative pharmacological target for their treatment.
Introduction
The RhoGTPases proteins regulate the actin cytoskeleton [1–3] and have emerged as key regulators of synaptic plasticity [4–6]. To evaluate the involvement of Rho GTPases on ageing processes, we pharmacologically modulated RhoGTPases pathways by administering in CD-1 male mice either fasudil, an inhibitor of the RhoA downstream effectors Rho kinases (ROCKs) [7] or CNF1, a bacterial toxin from Escherichia coli, known to activate in a constitutive manner the RhoGTPases [8].Aim
The aim of this work was to evaluate the effects of both ROCKs inhibition and RhoGTPases activation on cognitive and motor behavior and brain metabolism on a mouse model of healthy aging.Methods
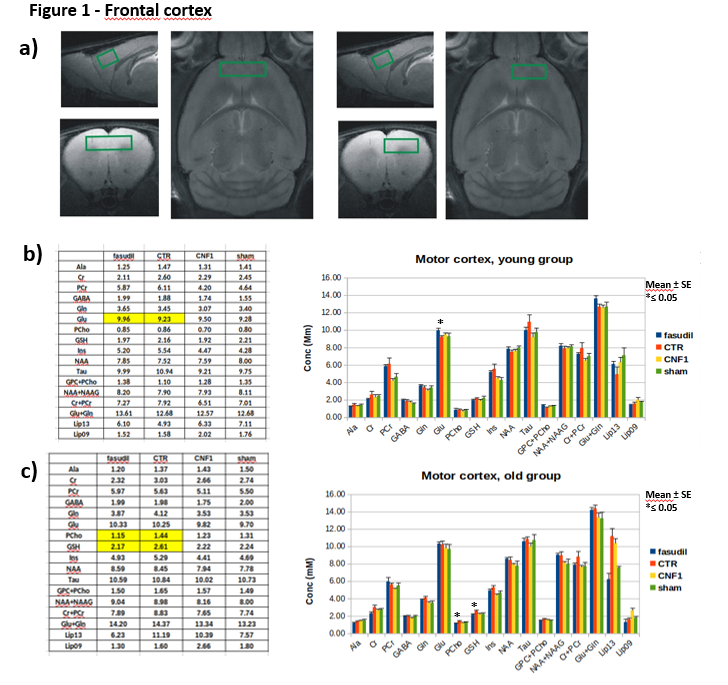
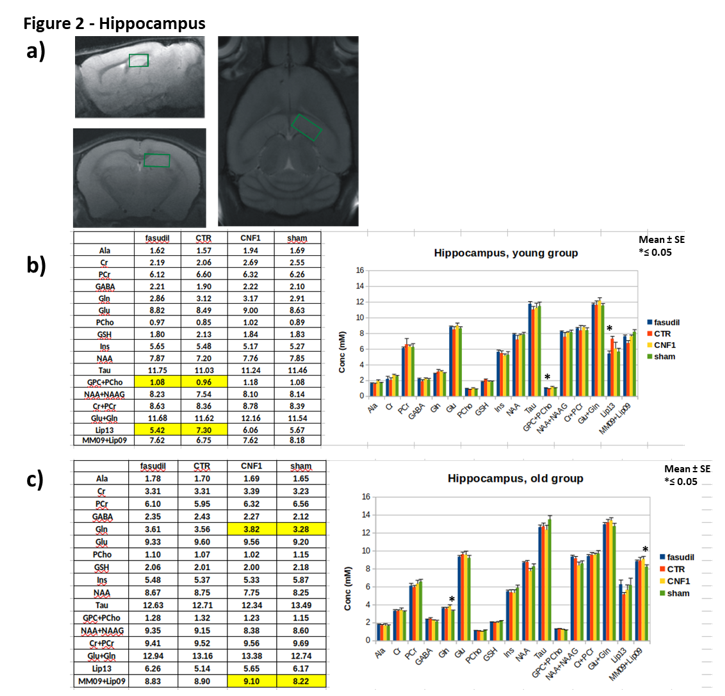
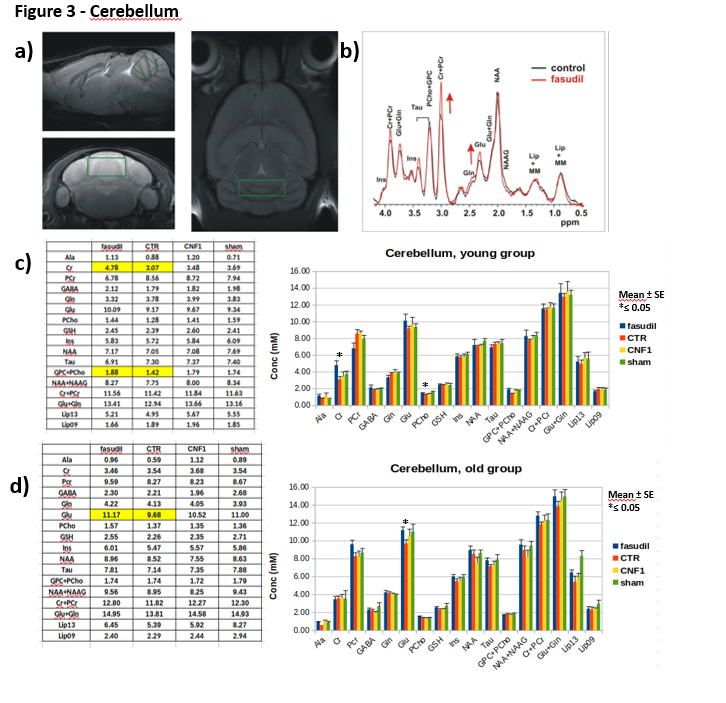
Male CD1 mice 1-month- and 15-month-old were administered with fasudil in drinking water (100mg/Kg/d) for three months and controls groups with fasudil-free (normal tap) water (n=12). At the age of 4months in the case of “young” group and of 18months for “old” group, animals were treated with 3 µl icv of a solution containing CNF1 toxin 1.34 x 10-8 M or the vehicle (n=12). Ten days after treatments, animals were tested with a battery of behavioral tests including Rotarod, open field and fear conditioning to assess motor coordination, locomotor activity/anxiety and cognitive performance, respectively. After behavioral tests, MRI/MRS experiments were performed on a reduced number of animals per group (6 for the young and 9 for the old group), on a small animal system (Pharmascan 7.0 T, Bruker) equipped with a cryo-probe. MRS was acquired by a quantitative protocol which includes a PRESS sequence, unsuppressed T2-corrected water signal as reference and LCModel fitting analysis [9]. MR spectra were collected from 3 regions: frontal cortex (PFC), right hippocampus (Hip) and cerebellum (Cer), as shown in Figure 1-3. These regions were selected for evaluating possible somatosensory (PFC) and cognitive (Hip) alterations and deficits in motor coordination (Cer). At the end of MRI/MRS, animals were euthanized and the same brain areas were collected for tissue analyses.Results
Aged mice showed decreased anxiety-related parameters respect to young mice, while no differences were found between the “young” and the “old” group in the case of motor parameters. There was no influence of the treatments in the motor coordination as shown by rotarod results. In the open field test both treatments increased locomotor activity and increased activity at the center of the arena, pointing to an overall anxiolytic effect in both young and aged mice.In fear conditioning test, during the context phase, fasudil and CNF1 treatments decreased % of freezing in young mice. Conversely, in the cued phase, CNF1-treated mice show a higher % of freezing respect to controls, in both young and old mice.
In the MR spectra of the “young” group, we detected an increase in glutamate concentration in the frontal cortex of fasudil-treated mice compared to their controls. Moreover, fasudil increases total choline content in all the examined regions and creatine/phosphocreatine ratio in the cerebellum. Finally, a decrease in the lipid signal (at 1.3 ppm) in the hippocampus was observed. No metabolic alterations due to CNF1 treatment was detected in all the examined regions.
In the “old” group we detected a decrease in PCho and GSH in the frontal cortex in the fasudil-treated mice compared to the controls. In the hippocampus, the CNF1-treated group had an increase in glutamine and lipids (0.9 ppm). In the cerebellum we measured an increase in glutamate in the fasudil-treated animals compared to the control group.
Tissue analysis are in progress.
Discussion and conclusions
Results showed that generally young mice present less anxiety and higher emotional memory response, compared to old animals. Moreover, ageing appears not to affect motor coordination and locomotion. The two Rho GTPases' modulators seem to act differently on the anxiety of old mice compared to young ones with an apparent greater anxiolytic effect limited to earlier age. Finally, CNF1 seems to favor cued fear memory at both ages. We observed also age- differences in the metabolism in response to both treatments. Fasudil increased total cholinecontent in all examined brain regions in the young mice and showed an influence on the choline metabolism in the aged mice. Conversely, only CNF1-treated old mice show an increase in hippocampal glutamine content.In conclusion, our data suggest a possible involvement of Rho GTPases in locomotion and in anxiety related to brain ageing. Moreover, we highlight a possible involvement of the GABAergic pathway leading to an improvement of synaptic plasticity and memory functions by targeting Rho GTPases.
Acknowledgements
Thanks are due to Andrea Martinelli (ISS) and Paolo Frassanito (ISS) for valuable animal care.References
1.Chardin, P.; Boquet, P.; Madaule, P.; Popoff, M.R.; Rubin, E.J.; Gill, D.M. The Mammalian G Protein RhoC Is ADP-Ribosylated by Clostridium Botulinum Exoenzyme C3 and Affects Actin Microfilaments in Vero Cells. EMBO J. 1989, 8, 1087–1092.
2.Hall, A. Rho GTPases and the Actin Cytoskeleton. Science 1998, 279, 509–514, doi:10.1126/SCIENCE.279.5350.509.
3.Paterson, H.F.; Self, A.J.; Garrett, M.D.; Just, I.; Aktories, K.; Hall, A. Microinjection of Recombinant P21(Rho) Induces Rapid Changes in Cell Morphology. J. Cell Biol. 1990, 111, 1001–1007, doi:10.1083/jcb.111.3.1001.
4.Govek, E.E.; Newey, S.E.; Van Aelst, L. The Role of the Rho GTPases in Neuronal Development. Genes Dev. 2005, 19, 1–49, doi:10.1101/gad.1256405.
5.Tolias, K.F.; Duman, J.G.; Um, K. Control of Synapse Development and Plasticity by Rho GTPase Regulatory Proteins. Prog. Neurobiol. 2011, 94, 133–148, doi:10.1016/j.pneurobio.2011.04.011.
6. Zhang, H.; Ben Zablah, Y.; Zhang, H.; Jia, Z. Rho Signaling in Synaptic Plasticity, Memory, and Brain Disorders. Front. cell Dev. Biol. 2021, 9, doi:10.3389/FCELL.2021.729076.
7. Yu, J.Z.; Chen, C.; Zhang, Q.; Zhao, Y.F.; Feng, L.; Zhang, H.F.; Meng, J.; Ma, C.G.; Xiao, B.G. Changes of Synapses in Experimental Autoimmune Encephalomyelitis by Using Fasudil. Wound Repair Regen. 2016, 24, 317–327, doi:10.1111/WRR.12407.
8. Boquet, P. The Cytotoxic Necrotizing Factor 1 (CNF1) from Escherichia Coli. Toxicon 2001, 39, 1673–1680, doi:10.1016/S0041-0101(01)00154-4.
9. Canese, R.; Pisanu, M.E.; Mezzanzanica, D.; Ricci, A.; Paris, L.; Bagnoli, M.; Valeri, B.; Spada, M.; Venditti, M.; Cesolini, A.; et al. Characterisation of in Vivo Ovarian Cancer Models by Quantitative 1H Magnetic Resonance Spectroscopy and Diffusion-Weighted Imaging. NMR Biomed. 2012, 25, 632–642, doi:10.1002/NBM.1779.
Figures