4124
Magnetic resonance elastography captures subtle changes in hippocampal CA1 stiffness between phases of the rat estrous cycle1Biomedical Engineering, University of Delaware, Newark, DE, United States, 2Psychological and Brain Sciences, University of Delaware, Newark, DE, United States, 3Psychology, University of Nebraska, Lincoln, NE, United States
Synopsis
Keywords: Small Animals, Elastography, sex differences, estrous, rat
Motivation: In rodent studies, the number of spines on hippocampal CA1 neurons decreases from proestrus to estrus, and complimentary reductions to hippocampal volume are observed in rodents and humans. It remains unknown if estrous-related changes to tissue microstructure alter the mechanical properties of CA1.
Goal(s): Evaluate the mechanical properties of CA1 between proestrus and estrus in the adult rat brain.
Approach: A novel rat Magnetic Resonance Elastography (MRE) protocol, measuring tissue deformation to determine stiffness and viscosity, was used to identify changes to the mechanical properties of CA1.
Results: CA1 stiffness increases between proestrus and estrus phases in female rats.
Impact: Results from this longitudinal study show that the stiffness of the hippocampal CA1 subregion increases between proestrus and estrus phases within-rat. We did not discover any changes to whole brain biomechanics between proestrus and estrus within-rat, consistent with the literature.
Introduction
Hippocampal microstructure changes across the estrous cycle in the adult female rat brain due to fluctuations in dendritic spine density on pyramidal neurons in the hippocampal CA1 subregion in response to varying hormone levels1. Rodent and human studies using traditional neuroimaging techniques have uncovered that the volume of CA1 fluctuates across the estrous/menstrual cycle2,3, and hippocampal CA1 in the female brain maintains high neuroplastic potential in adulthood.Magnetic Resonance Elastography (MRE) is a neuroimaging technique that is sensitive to tissue mechanical properties reflecting microstructure composition and organization, demonstrating an added (and/or complimentary) value of MRE relative to conventional MRI protocols. MRE produces spatially-resolved maps of brain tissue mechanical properties by imaging small tissue deformations and estimating material parameters, such as stiffness and viscosity, via inverse solution of the underlying equations of motion. MRE has successfully measured neuroplasticity-related changes to whole brain and hippocampal microstructure in the adult brain4,5. It remains unknown if MRE can capture subtle alterations to hippocampal microstructure that occur over brief, routine neuroendocrinological cycles.
This longitudinal study uses a rat MRE protocol to evaluate changes in CA1 microstructure across the estrous cycle. In vivo MRE results will be verified with histological quantification of dendritic spines1 and astrocytic processes6. These data address the remaining gap in our knowledge surrounding the potential for sex differences in hippocampal mechanical properties, and understanding these differences may inform studies assessing progression of neurological disorders with greater prevalence in females (i.e., Alzheimer's Disease, Multiple Sclerosis, and catamenial epilepsy).
Methods
Six adult female Long-Evans rats with a regular 4-day estrous cycle were used in the study. MRE scans during each of the four cycle phases were performed (Figure 1). MRE data were acquired with a custom echoplanar imaging sequence with a 9.4T Bruker Biospec scanner. A piezoelectric actuator was attached to a custom carbon fiber bite bar to deliver vibrations at 700 Hz to anesthetized rats maintained on 1-3% isoflurane in oxygen and at 32-36 ℃ for a total 1-hour scan protocol. MRE imaging parameters included: TE/TR = 60/4000 ms; 20x20 mm FOV; 80x80 matrix; 40 slices, 0.5 mm thick; 0.25x0.25x0.50 mm resolution. Images with OSS-SNR < 3 were discarded7 and the rat was rescanned at the same phase. Maps of mechanical properties, including stiffness and damping ratio, were estimated from displacement images with nonlinear inversion (NLI) formulated without boundary conditions8,9. We predicted that CA1 stiffness would be reduced between proestrus and estrus, mirroring the phase-specific reduction in dendritic spine density in this region.Results
We performed within-rat, repeated measures ANOVA on the longitudinal stiffness and damping ratio values from the whole brain and the hippocampal CA1 subregion between the proestrus and estrus cycle phases. CA1 stiffness increased from proestrus to estrus (F1,5 = 7.21, p = 0.04, ηp2 = 0.60; Figures 2 & 3), in contradiction to our original hypothesis. The damping ratio of CA1 tissue did not change significantly between the phases (p > 0.05). Consistent with the rodent and human literature, we did not observe significant changes to whole brain stiffness or damping ratio between proestrus and estrus (p > 0.05; Table 1).Conclusions
We show that estrous phase has a significant effect on the hippocampal tissue stiffness in the female rat brain. We anticipated that CA1 would soften between proestrus and estrus, in parallel to the known reduction to dendritic spine number and volume of CA1. However, our preliminary findings suggest that CA1 stiffens in estrus. Smith et al. demonstrated using atomic force microscopy that newly formed, immature dendritic spines are extremely “soft”10, while the stiffest spines are mature spines with axonal connections. It is well-established that the population of spines that are produced and retracted across the estrous cycle seldom creates lasting axonal contacts. Thus, the reduction of these highly plastic, immature spines from proestrus to estrus could lead to a stiffening of the CA1 as fewer “soft” spines are present in CA1 during estrus. Ultimately, our preliminary findings show that rat MRE is a valuable tool for assessing tissue microstructure plasticity that occurs within 24 hours, which could impact hippocampal-dependent memory function and onset of certain disease symptoms. Moreover, estrous cycle may confound MRE measurements of hippocampal stiffness in the female brain.Acknowledgements
NIH/NIBIB R01 EB027577 (Johnson), NIH 2P20GM10365 (University of Delaware Center for Biomedical and Brain Imaging), NIH/NIAAA R01 AA027269-01 (Klintsova)
References
[1] Woolley et al. Naturally occurring fluctuation in dendritic spine density on adult hippocampal pyramidal neurons. The Journal of Neuroscience, 1990; 10(12), 4035–4039.
[2] Zsido et al. Ultra-high-field 7T MRI reveals changes in human medial temporal lobe volume in female adults during menstrual cycle. Nature Mental Health, 2023; 1(10), 761–771.
[3] Qiu et al. Hippocampal volumes differ across the mouse estrous cycle, can change within 24 hours, and associate with cognitive strategies. NeuroImage, 2013; 83, 593–598.
[4] Schwarb et al. Aerobic fitness, hippocampal viscoelasticity, and relational memory performance. NeuroImage, 2017; 153, 179–188.
[5] Sandroff et al. Exercise training effects on memory and hippocampal viscoelasticity in multiple sclerosis: a novel application of magnetic resonance elastography. Neuroradiology, 2017; 59(1), 61-67
[6] Klintsova et al. Astrocytic volume fluctuates in the hippocampal CA1 region across the estrous cycle. Brain Research, 1995; 690(2), 269–274.
[7] McGarry et al. An octahedral shear strain based measure of SNR for 3D MR elastography. Physics in Medicine and Biology, 2011; 56(13), N153-N164.
[8] McGarry et al. Multiresolution MR elastography using nonlinear inversion. Medical Physics. 2012; 39(10):6388-6396.
[9] Kurtz et al. Minimizing measurement-induced errors in viscoelastic MR elastography. IEEE Transactions on Medical Imaging. 2023; doi: 10.1109/TMI.2023.3329293.
[10] Smith et al. Dendritic Spine Viscoelasticity and Soft-Glassy Nature: Balancing Dynamic Remodeling with Structural Stability. Biophysical Journal, 2007; 92, 1419–1430.
Figures
Figure 1. Experimental timeline depicting the timing of facility acclimation and longitudinal MRE scanning across the estrous cycle.
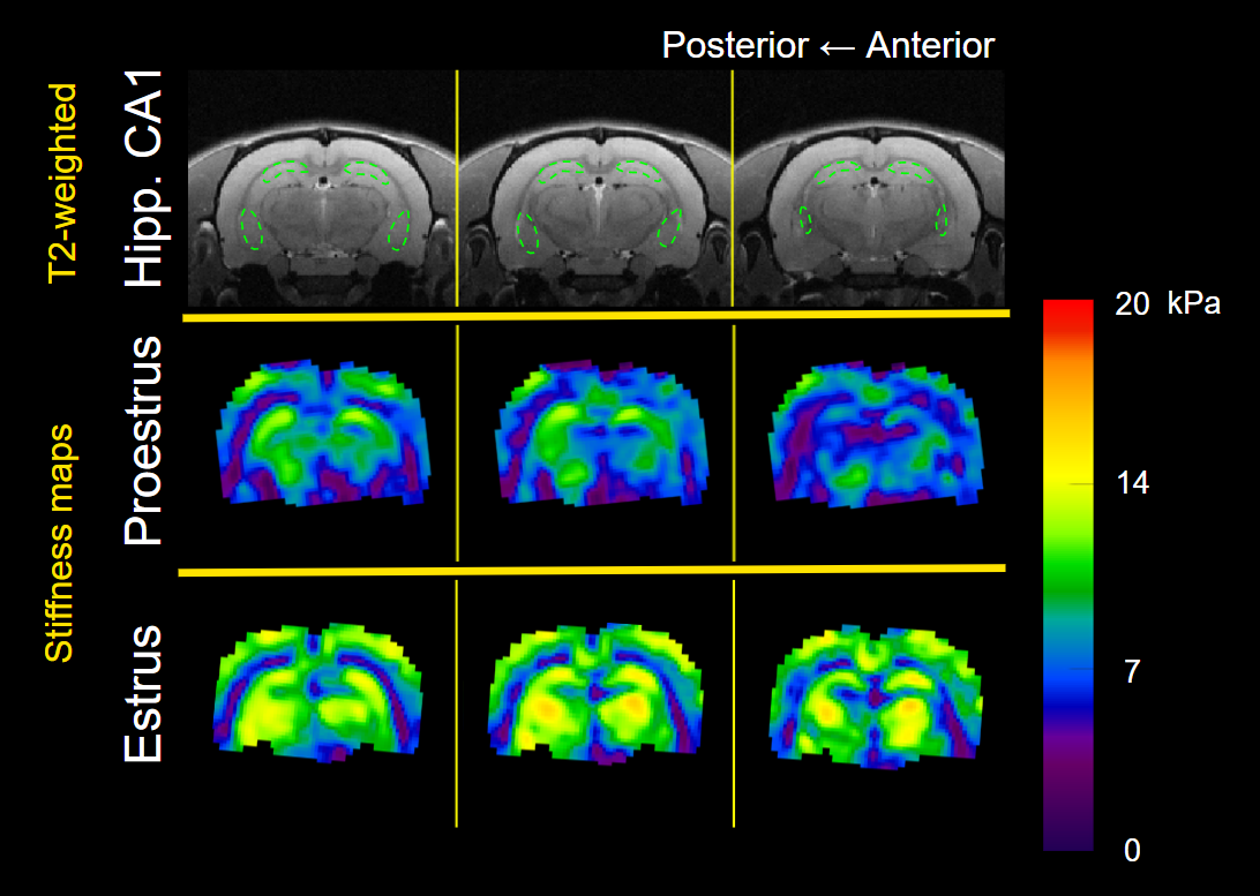
Figure 2. Representative shear stiffness (μ) maps containing hippocampal CA1 during the proestrus and estrus cycle phases. T2-weighted anatomical scan illustrates the position of the ROI for CA1.
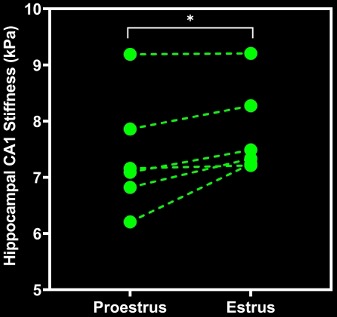
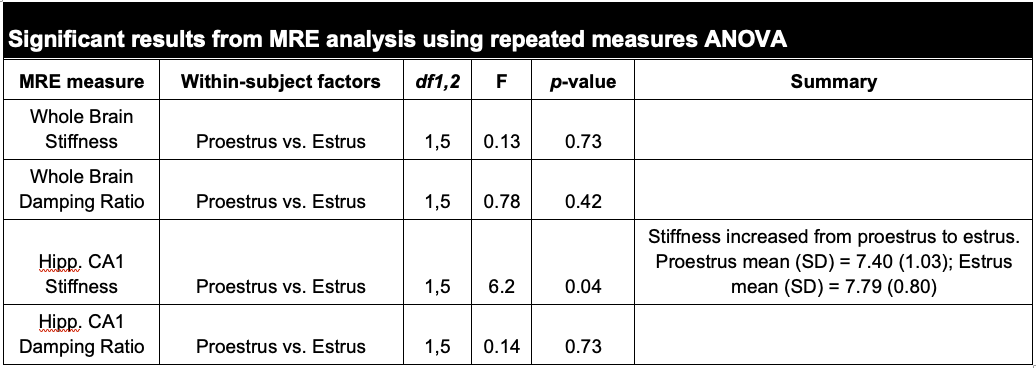
Table 1. Summary of MRE property results. Repeated measures ANOVA analysis revealed a significant stiffening of the CA1 subregion of the hippocampus from proestrus to estrus phases.