4036
Exploring the Effects of Phospholipase A2 Inhibition on Brain Metabolism and Pathology in a Rat Model of Alzheimer’s Disease1Medical Biophysics, University of Toronto, Toronto, ON, Canada, 2Sunnybrook Research Institute, Toronto, ON, Canada, 3Laboratory Medicine and Pathobiology, University of Toronto, Toronto, ON, Canada
Synopsis
Keywords: Alzheimer's Disease, Spectroscopy
Motivation: Current AD treatments do not improve cognitive deficits. Gaining a deeper understanding of the mechanisms underlying AD may reveal new therapeutic targets.
Goal(s): Previous work demonstrated elevated choline levels in a preclinical model which may implicate the PLA2 pathway in AD. We aim to explore the role of PLA2 in AD.
Approach: To understand the role of PLA2 in AD, we pharmacologically inhibited PLA2 in a preclinical model of AD. Metabolic, behavioural, and pathological aspects of AD were then explored using MRS, Barnes maze, immunostaining, and RNA sequencing.
Results: Preliminary data indicate PLA2 inhibition reduces choline levels and improves long-term memory formation.
Impact: This work may reveal PLA2 as new therapeutic target that improves cognitive deficits in Alzheimer’s disease. By linking behaviour, pathology, and metabolic processes, this study will give a well-rounded picture of how PLA2 activity impacts AD progression.
Introduction
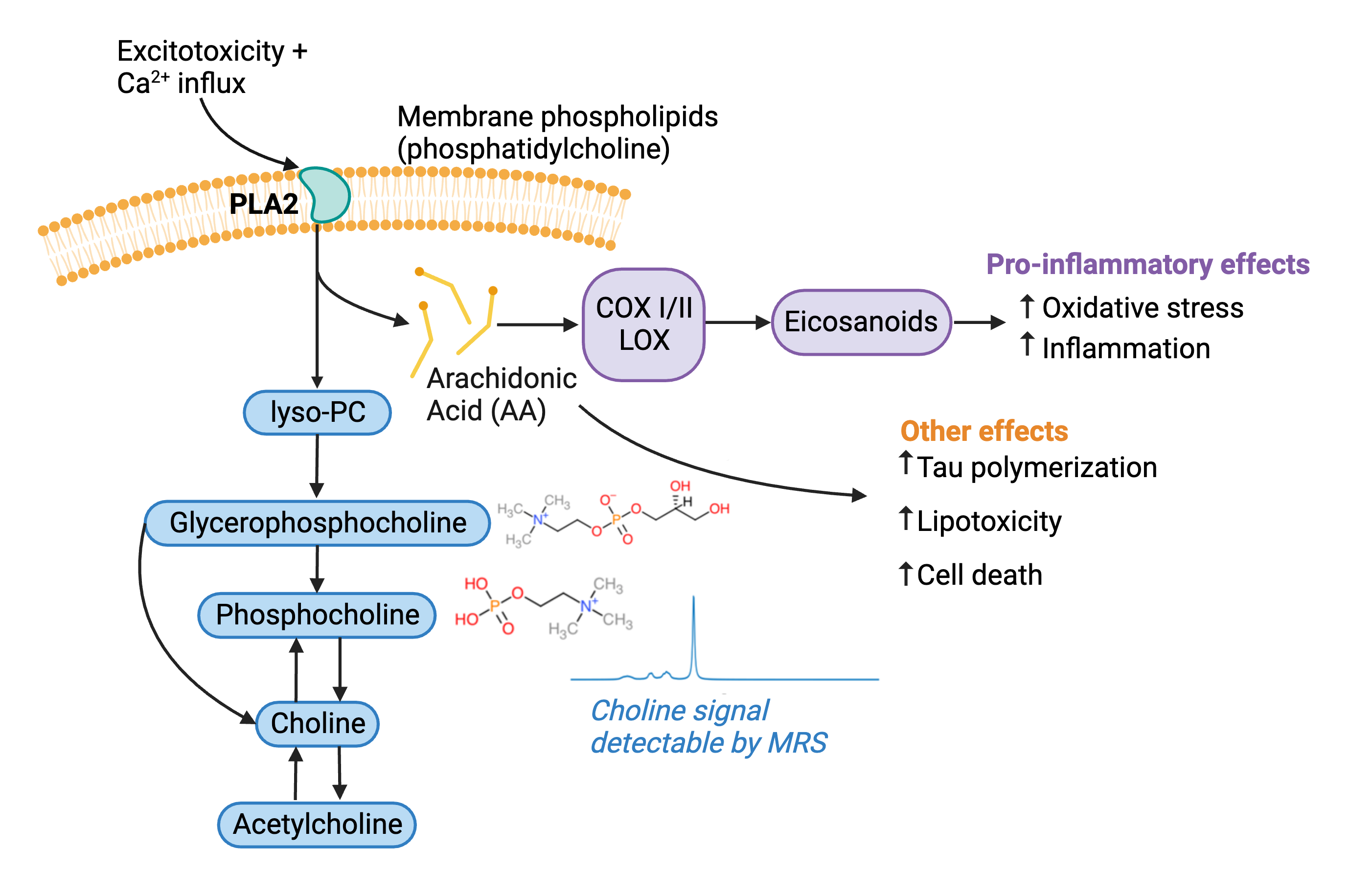
Alzheimer’s disease (AD) is a devastating neurodegenerative disorder which has no effective treatment options to delay or halt disease progression1. The main characteristics of this disorder are amyloid-β plaques, tau neurofibrillary tangles, neuronal atrophy, and cognitive deficits2. Some recent treatment options have focused on reducing amyloid-β plaques but have failed in phase 3 clinical trials1. As a result, it is necessary to explore alternate treatment options and target different pathways affected by AD. Magnetic resonance spectroscopy (MRS) offers a non-invasive modality to explore disease pathogenesis and treatment efficacy. Previous MRS studies in both animal models3,4 and humans5-8 have shown an increase in choline in AD. The breakdown of membrane phosphatidylcholine by the enzyme phospholipase A2 (PLA2) generates glycerophosphocholine and phosphocholine which are measurable by MRS9. Thus, elevated PLA2 activity may play a role in AD progression, illustrated by the working model in figure 1. This study aims to gain a deeper understanding of the role of PLA2 in AD through the simultaneous observation neurochemistry, behaviour, and pathology following PLA2 inhibition in an animal model of AD.Methods
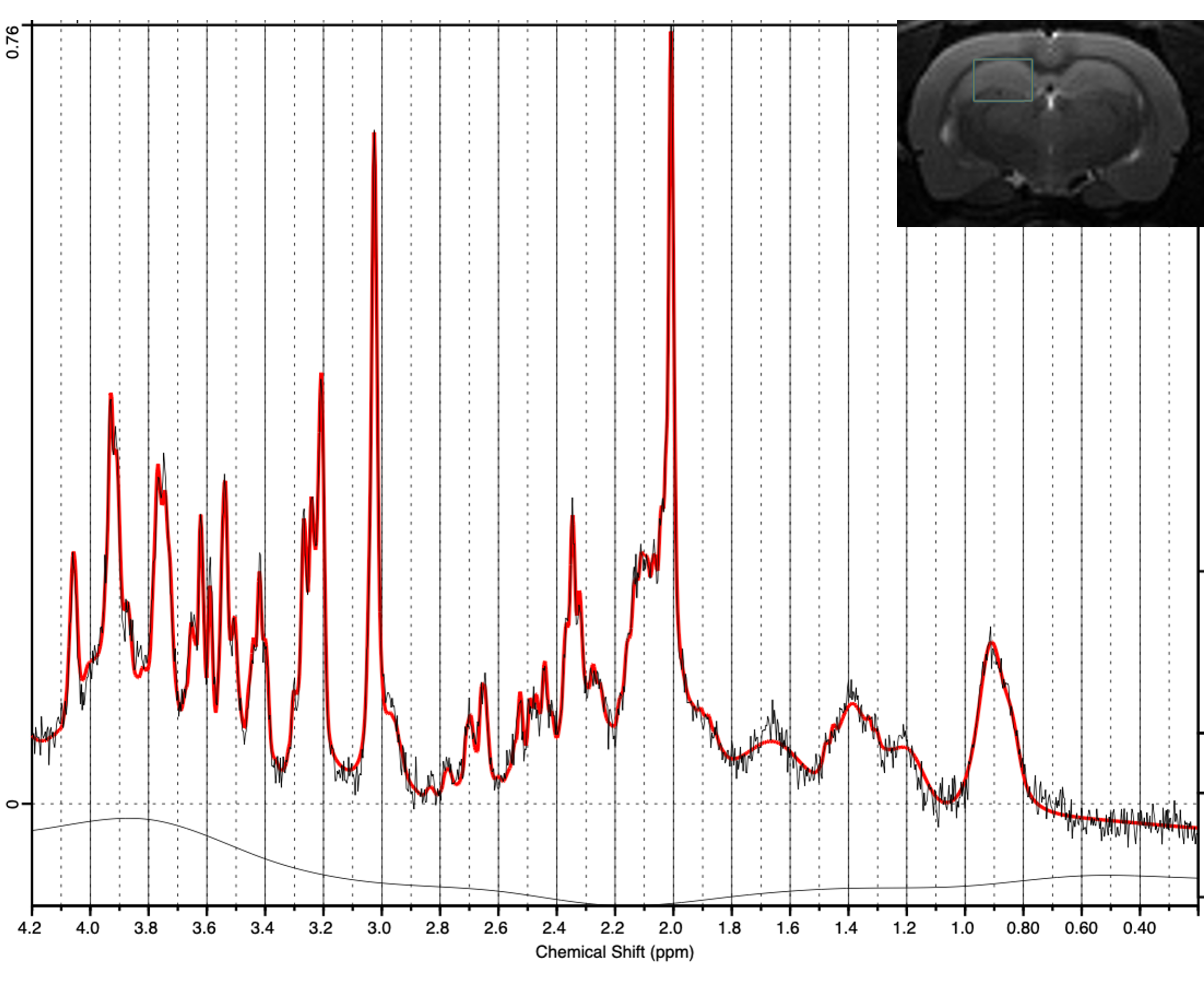
This study used the TgF344-AD rat model at approximately 13 months because previous research has demonstrated significantly elevated choline levels at this age3. This model is unique in that it exhibits a complete range of AD pathology10. The global PLA2 inhibitor mepacrine was used because it crosses the blood brain barrier and has been shown to inhibit PLA2 in the brain11-13. Brain metabolite concentrations, specifically total choline (TCh) levels were explored using MRS in both transgenic (Tg, n=8) and non-transgenic (NTg, n=6) animals before and after one week of treatment with either saline (n=7) or mepacrine (n=7). Proton MRS was acquired on a 7 Tesla MRI system (Bruker, Billerica MA, USA) from the right dorsal hippocampus (Figure 2) using a PRESS sequence (TR/TE = 2500/8.8 ms, 256 averages). After the follow-up MRS scan, animals completed the Barnes maze. Briefly, animals were required to find an escape hole on a platform with 19 other false escape holes. Animals completed acquisition training, a probe trial, and reversal training in order to test long-term memory formation and cognitive flexibility. To explore pathological changes, animals were sacrificed following completion of the Barnes maze. Immunohistochemistry and immunofluorescence will be completed to observe amyloid-β (6F3D), tau (PHF1), and inflammation (microglia (IBA1), astrocytes (GFAP)). Other tissue will be collected for RNA sequencing to explore changes in genetic expression following PLA2 inhibition.Results
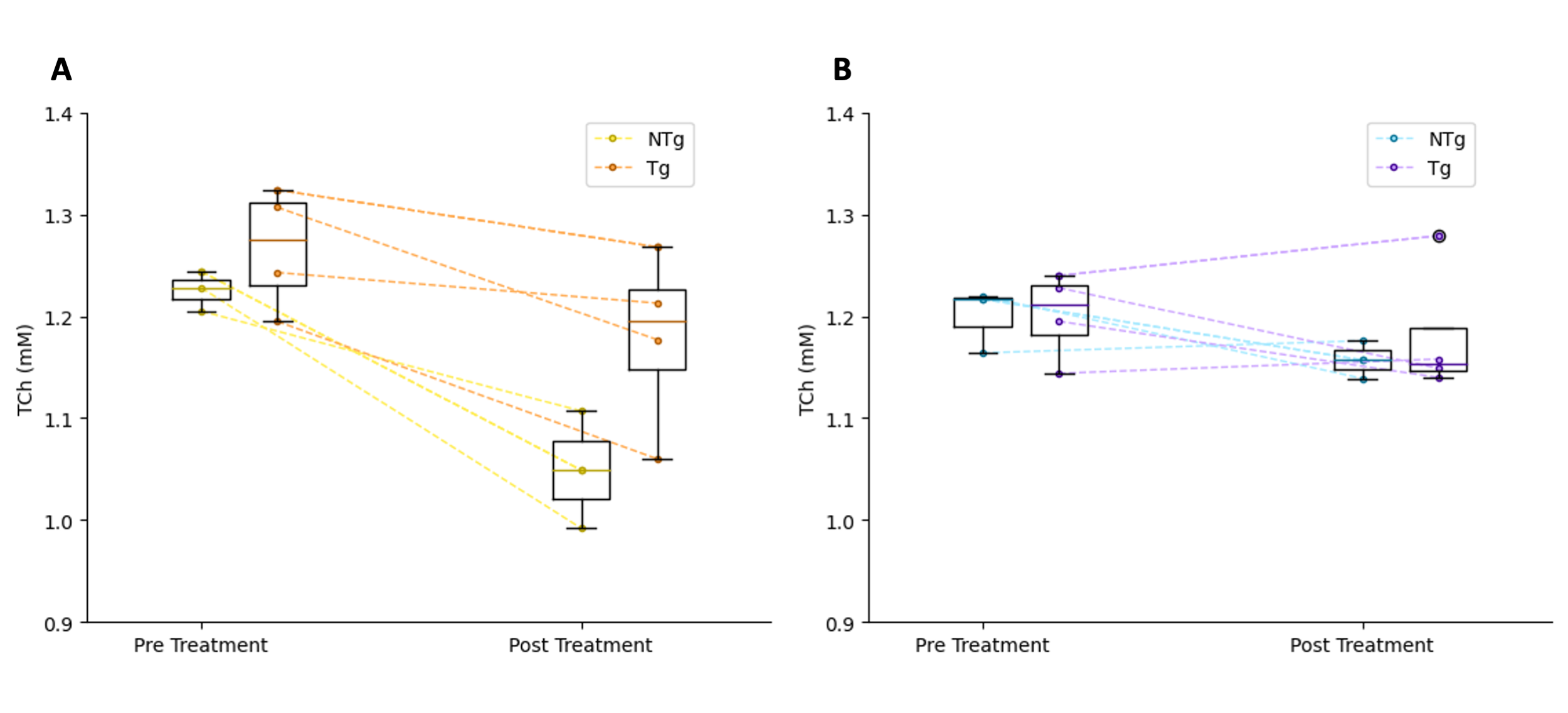
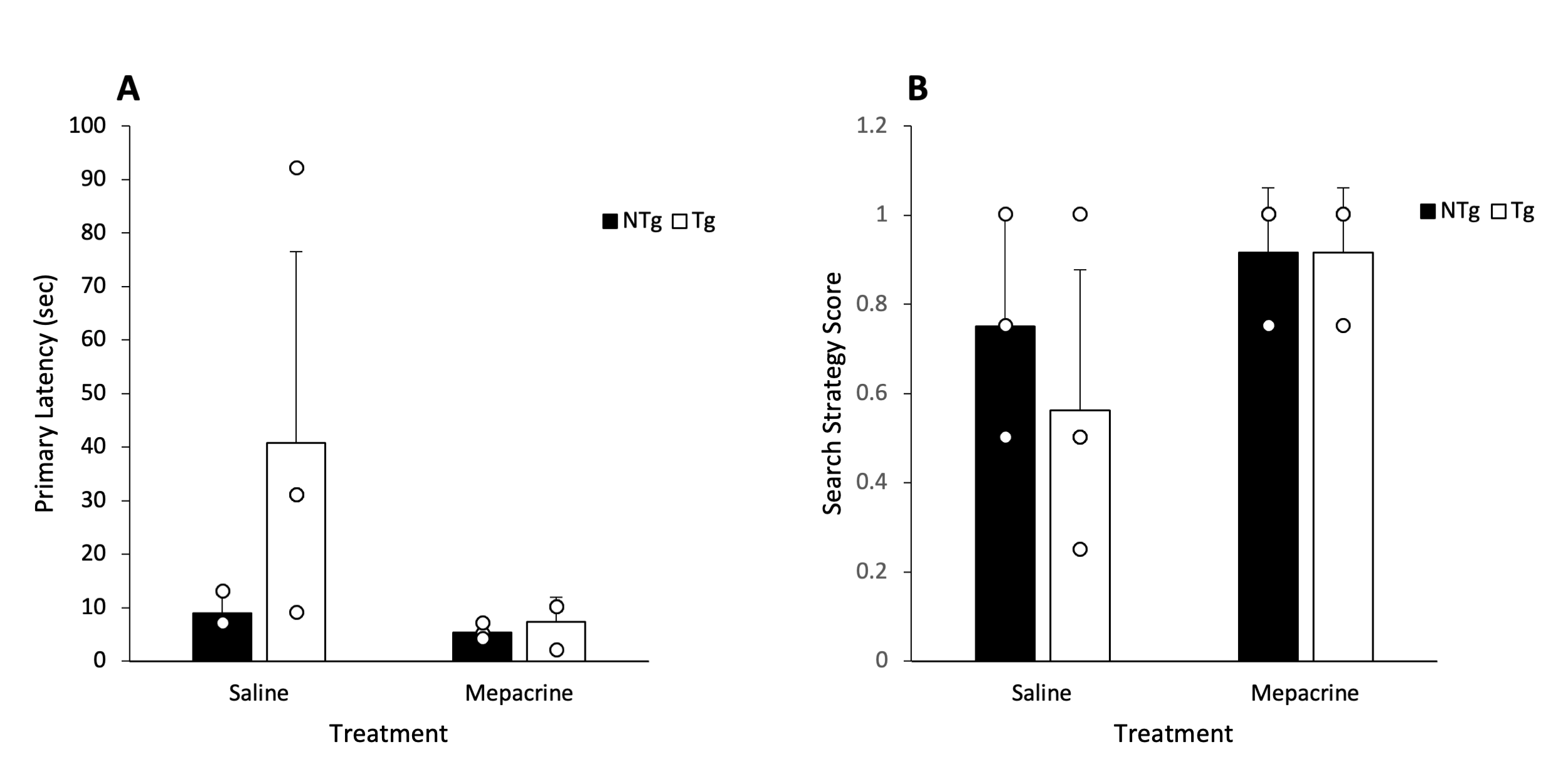
TCh levels were observed to decrease in the hippocampus following one week of treatment with mepacrine (Figure 3A). TCh levels were steadier after one week in the saline controls (Figure 3B). Contrary to our expectations, TCh levels at baseline did not appear to differ between Tg and NTg genotypes. In the probe trial of the Barnes maze, Tg animals treated with mepacrine were observed to have a similar primary latency to the escape hole as NTg controls (Figure 4A). Animals treated with mepacrine had a search strategy score closer to 1, demonstrating a more direct search strategy (Figure 4B). NTg animals treated with mepacrine appear to have similar primary latency and search strategies when compared to NTg animals treated with saline.Discussion
Although higher levels of baseline TCh have not yet been observed in Tg animals in this study, previous studies have demonstrated an overlap in TCh between Tg and NTg animals3. Therefore, as we add more animals to this study, we expect to see this pattern start to emerge. The decrease in TCh in mepacrine treated animals shows promise that mepacrine is effectively inhibiting PLA2.The preliminary Barnes maze results show promise that PLA2 inhibition improves long-term spatial memory formation. This agrees with a previous study that demonstrated genetic ablation of PLA2 improved long-term spatial memory formation in a mouse model of AD14.Conclusion
The relevance of the present study is that it may reveal a potential mechanism underlying AD pathology. Although the data collected are not complete, we have already observed the inhibition of PLA2 demonstrated by decreased choline in MRS and improved long-term memory formation in treated animals. We expect this pattern to continue as more data are collected. In addition, exploring pathological changes through immunostaining and RNA sequencing may reveal mechanisms underlying the behavioural changes caused by PLA2 inhibition. Overall, we have shown that PLA2 expression in AD may contribute to memory impairment. This demonstrates promise in identifying a therapeutic target which may improve cognitive deficits in AD.Acknowledgements
This work is supported by the Canadian Institutes for Health Research (JN Grant #: PJT-165869) and the National Sciences and Engineering Research Council (JN Grant #: RGPIN-2020-05917).References
1. Yiannopoulou KG, Papageorgiou SG. Current and Future Treatments in Alzheimer Disease: An Update. J Cent Nerv Syst Dis. 2020;12:1179573520907397.
2. Terry RD, Masliah E, Salmon DP, et al. Physical basis of cognitive alterations in alzheimer's disease: Synapse loss is the major correlate of cognitive impairment. Ann Neuol. 1991;30(4):572-580.
3. Fowler CF, Goerzen D, Devenyi GA, et al. Neurochemical and cognitive changes precede structural abnormalities in the TgF344-AD rat model. Brain Comm 2022;4(2):1-16.
4. Choi JK, Carreras I, Aytan N, et al. The effects of aging, housing and ibuprofen treatment on brain neuroschiemistry in a triple transgene Alzheimer’s disease mouse model using magnetic resonance spectroscopy and imaging. Brain Res. 2014;1590:85-96.
5. Marhanska M, McCarten JR, Hodges JS, et al. Distinctive Neurochemistry in Alzheimer’s Disease via 7T In Vivo Magnetic Resonance Spectroscopy. J Alzheimers Dis. 2019;68(2):559-569.
6. Joe E, Medina LD, Ringman JM, et al. 1H MRS Spectroscopy in Preclinical Autosomal Dominant Alzheimer Disease. Brain Imaging Behav. 2019;13(4):925-932.
7. Kantarci K, Petersen RC, Boeve BF, et al. 1H MR SPECTROSCOPY IN COMMON DEMENTIAS. Neurology. 2004;63(8):1393-1398.
8. Meyerhoff DJ, MacKay S, Constans JM, et al. Axonal injury and membrane alterations in Alzheimer's disease suggested by in vivo proton magnetic resonance spectroscopic imaging. Ann Neurol. 1994;36(1):40-47.
9. Klein J. Membrane breakdown in acute and chronic neurodegeneration: focus on choline-containing phospholipids. J Neural Transm 2000;107:1027-1063.
10. Cohen RM, Rezai-Zadeh K, Weitz TM, et al. A transgenic Alzheimer rat with plaques, tau pathology, behavioral impairment, oligomeric aβ, and frank neuronal loss. J Neurosci 2013;33(15):6245-6256.
11. Ong WY, Lu XR, Ong BKC, et al. Quinacrine abolishes increases in cytoplasmic phospholipase A2 mRNA levels in the rat hippocampus after kainite-induced neuronal injury. Exp Brain Res 2003;148:521-524.
12. Ong WY, Lu XR, Horrocks LA, et al. Induction of astrocytic cytoplasmic phospholipase A2 and neuronal death after intracerebroventribular carrageenan injection, and neuroprotective effects of quinacrine. Exp Neurol 2003;183:449-457.
13. Kencebay C, Derin N, Ozsoy O, et al. Merit of quinacrine in the decrease of ingested sulfite-induced toxic action in rat brain. Food Chem Tox 2013;52:129-136.
14. Sanchez-Mejia RO, Newman JW, Toh S, et al. Phospholipase A2 reduction ameliorates cognitive deficits in a mouse model of Alzheimer’s disease. Nat Neurosci 2008;11:1311-1318.
Figures