3901
Insights into neurodegeneration in Alzheimer’s disease from regional Aβ aggregation, iron level, and gene expression in postmortem human brain1Center for Brain Imaging Science and Technology, Zhejiang University, Hangzhou, China, 2College of Biomedical Engineering and Instrument Science, Zhejiang University, Hangzhou, China, 3School of Biomedical Engineering, Shanghai Jiao Tong University, Shanghai, China, 4Stanford University Graduate School of Education, Department of Radiology, Standford University, Stanford, CA, United States, 5National Human Brain Bank for Health and Disease, School of Brain Science and Brain Medicine, Zhejiang University, Hangzhou, China, 6Department of Imaging Sciences, University of Rochester, Rochester, NY, United States, 7School of Physics, Zhejiang University, Hangzhou, China, 8State Key Laboratory of Brain-Machine Intelligence, Zhejiang University, Hangzhou, China
Synopsis
Keywords: Alzheimer's Disease, Alzheimer's Disease
Motivation: Quantifying Aβ in patients with Alzheimer's disease poses a challenge due to the colocalization of Aβ accumulation and iron deposition.
Goal(s): Our goal was to simultaneously quantify Aβ and iron in ex-vivo human brains affected by AD.
Approach: We used a novel subvoxel QSM method to measure Aβ and iron levels. The gene transcriptomic profiles were further investigated using PLS and ontological analysis.
Results: Regions with higher diamagnetic and paramagnetic susceptibility were found higher levels of gene expression relating to the protein modification process and metal ion binding, as well as a relative abundance of exCA and glutamatergic neurons.
Impact: The quantification of diamagnetic and paramagnetic susceptibility via APART-QSM can offer valuable insights into regional-specific vulnerabilities in Alzheimer’s disease, particularly those related to Aβ aggregation and iron accumulation. This can aid clinicians to better find therapeutic targets.
Introduction
The formation of amyloid plaques through the aggregation of beta amyloid (Aβ) is one of the hallmarks of Alzheimer's disease (AD)1. Concurrently, the accumulation of iron has long been suspected as a fundamental cause of AD, as it interacts with Aβ through metal binding and likely contributes to the pathological processes of the disease2,3. However, it is challenging to accurately quantify iron levels using MRI due to the presence of Aβ. Recent studies showed Aβ is diamagnetic and can generate strong contrast on susceptibility maps4–6. In this study, we utilized the iterative magnetic susceptibility sources separation (APART-QSM) method to simultaneously distinguish diamagnetic and paramagnetic susceptibility sources7, exploring Aβ aggregation and brain iron levels in ex-vivo human brains from individuals with AD and healthy controls (HC). Additionally, we examined the biological processes and cell types associated with this transcriptomic profile. Our findings provide further mechanistic insights into regional selective vulnerabilities in AD, specifically those related to Aβ accumulation and iron deposition.Methods
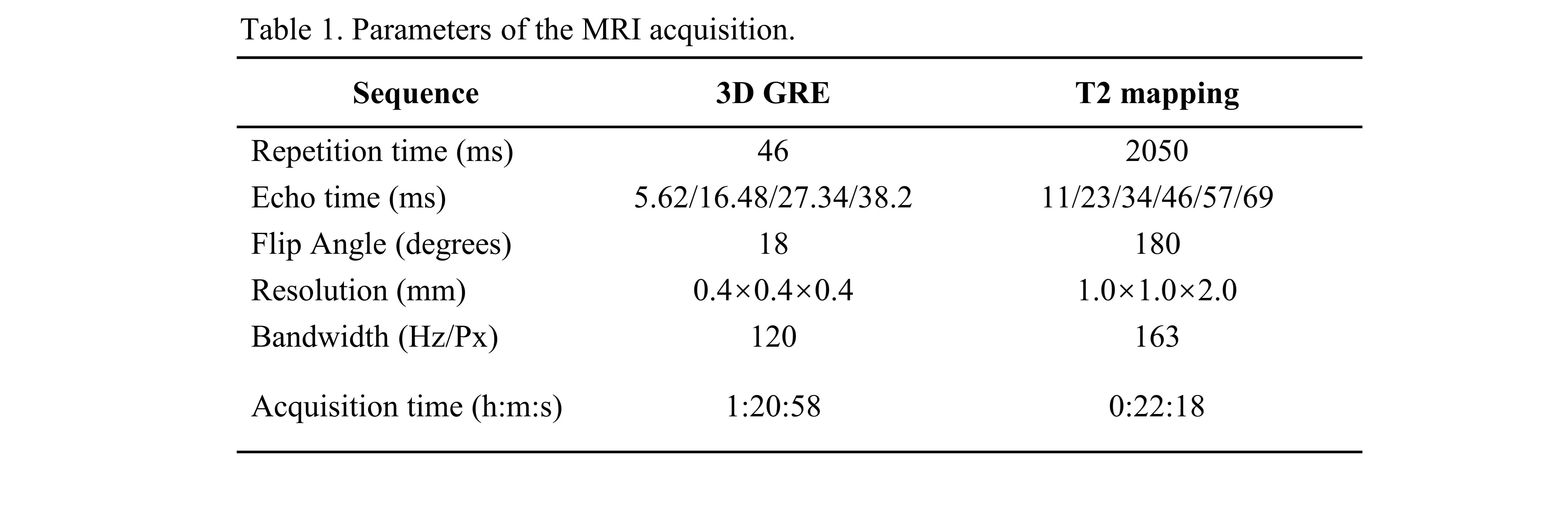
In this study, four right hemispheres of formalin-fixed human brain (2 AD, 2 HC) were obtained from the National Human Brain Bank for Health and Disease in Zhejiang University, China. Postmortem brains were processed as detailed in a previous study8. The MRI data were acquired on a 7T MAGNETOM scanner using a NOVA head-neck coil, with the main parameters listed in Table 1. Diamagnetic (χdia) and paramagnetic (χpara) susceptibility were computed using APART-QSM algorithm7. Regions of interest (ROI) were delineated using the multi-modal parcellation (MMP) atlas9,10, mean values in each ROI were extracted for all samples. Seven ROIs were excluded due to signal inhomogeneity.The association between the difference map of χdia and χpara in AD versus HC and gene expression was investigated using PLS regression11. Gene expression data was obtained from the AIBS and projected onto the MMP Atlas12,13. Gene ontological (GO) enrichment analysis was performed on the significantly positively weighted genes identified by PLS214. To investigate the cell types associated with those highly weighted genes, an expression-weighted cell-type enrichment (EWCE) analysis was conducted15, using the AIBS single-cell transcription dataset and a human derived dataset (DroNc-human) 16,17.
Results & Discussion
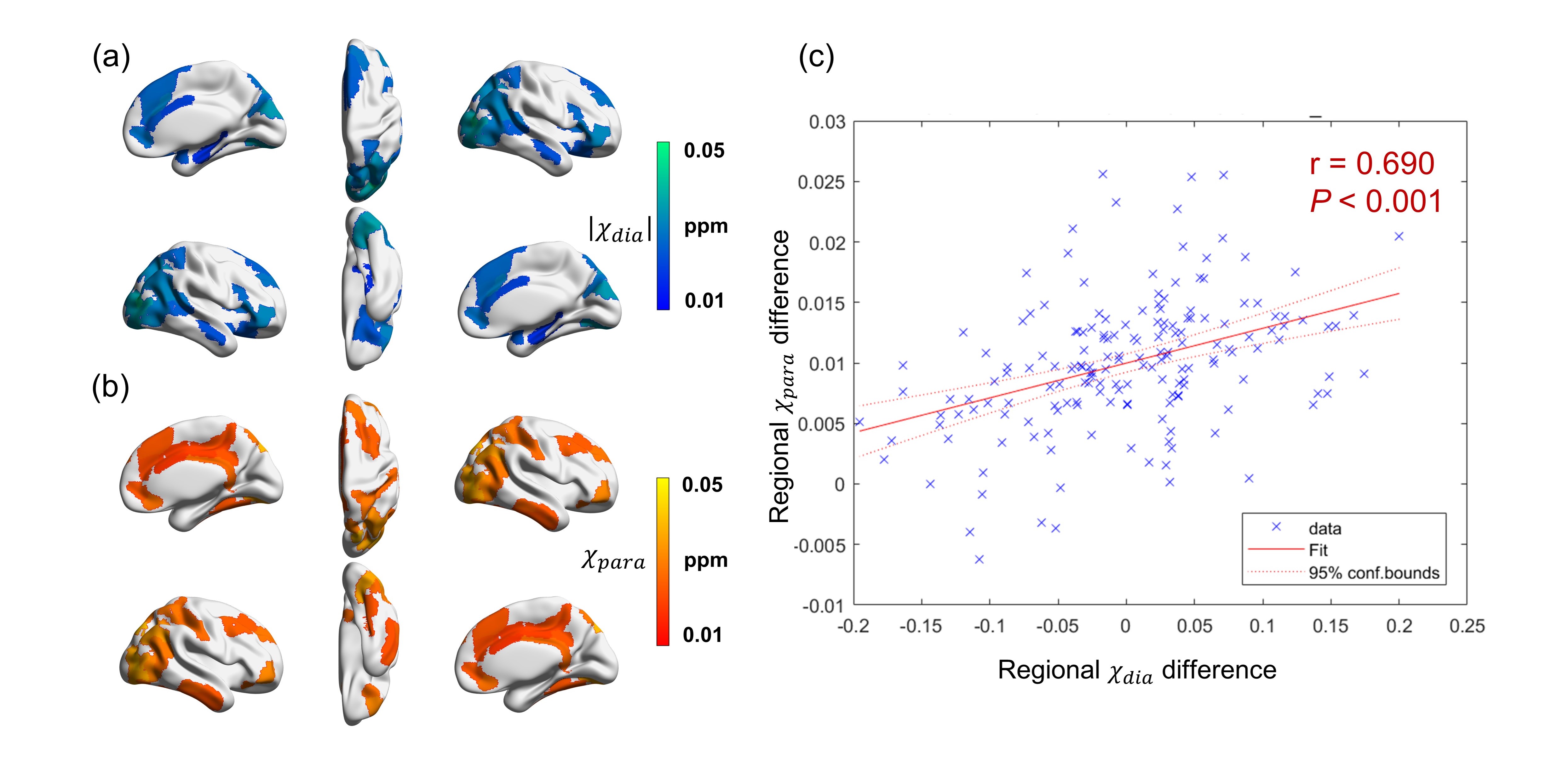
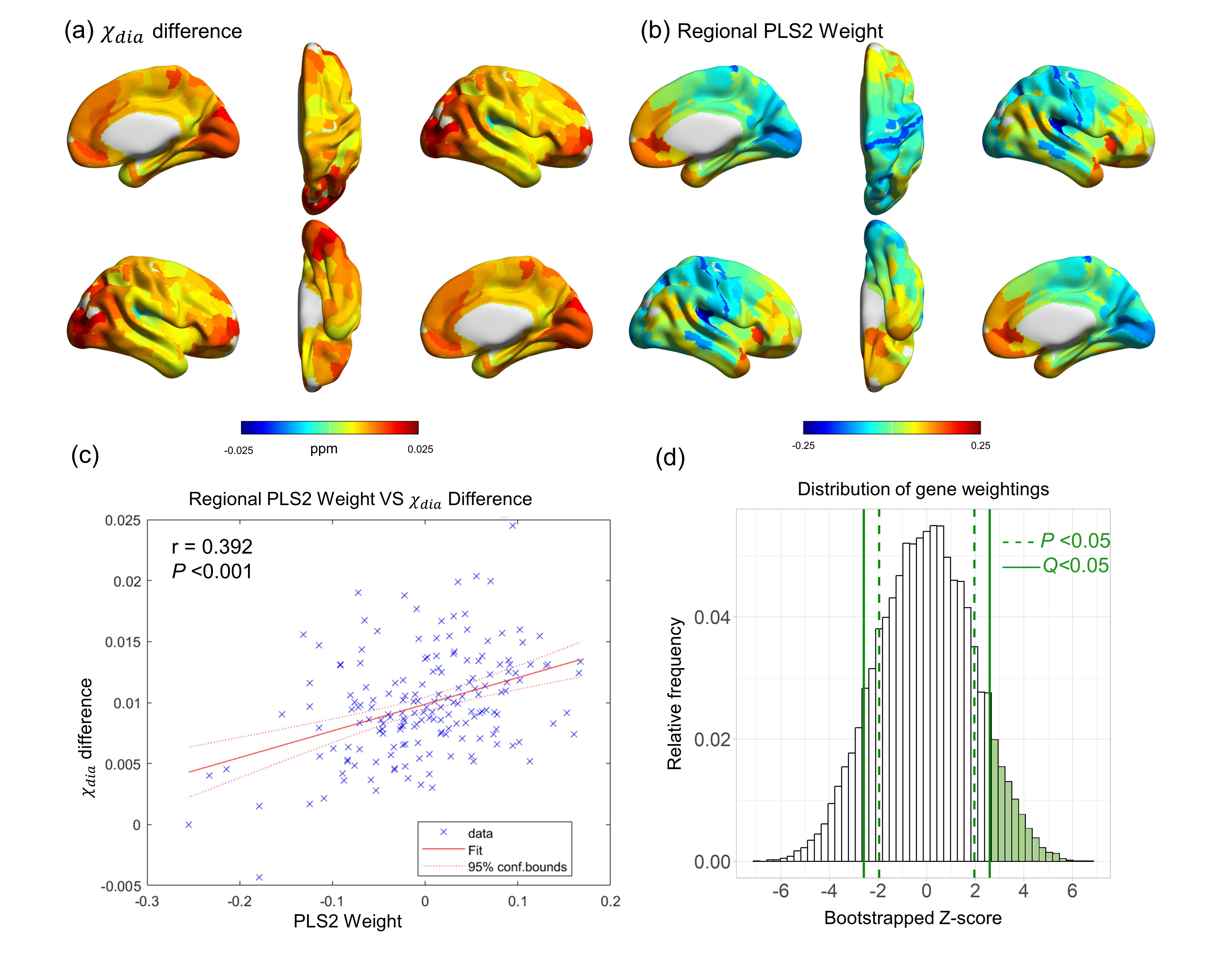
According to the pathological diagnosis of individuals with AD18, ex-vivo brains display positive Aβ staining in regions including the middle frontal gyrus, posterior hippocampus, parietal lobe, and occipital lobe. In those regions, significant increases of χdia values were also detected (Fig. 1a). Moreover, a substantial overlapping is observed between χdia and χpara in regions characterized by significant distinctions between AD and HC. Notably, a significant positive correlation was observed between the regional χdia and χpara difference (Fig. 1c), indicating a colocalization of Aβ aggregation and iron deposition in the AD brain19.The difference map of χdia between AD and HC had a similar spatial pattern to the regional linearly weighted sum of gene expression scores defined by the PLS2 (Fig. 2). The PLS2 explained the most variance in the difference map. The PLS2 gene expression weights showed a strong positive correlation with the difference between AD and HC in χdia (Fig. 2c), suggesting that genes that were positively weighted on PLS2 were also more highly expressed in cortical brain regions with higher χdia.
Using GO analyses, we found sets of biological processes associated with upweighted genes (Table 2). Genes more highly expressed in regions with higher χdia and χpara values in AD were both enriched for GO terms relating to the protein modification process, post-translational protein modification, cellular response to stress, which are associated with Aβ aggregation20. In addition, the GO items in χpara analysis also involved metal ion binding (P=2.16×10-8), which can be associated with the iron accumulation21.
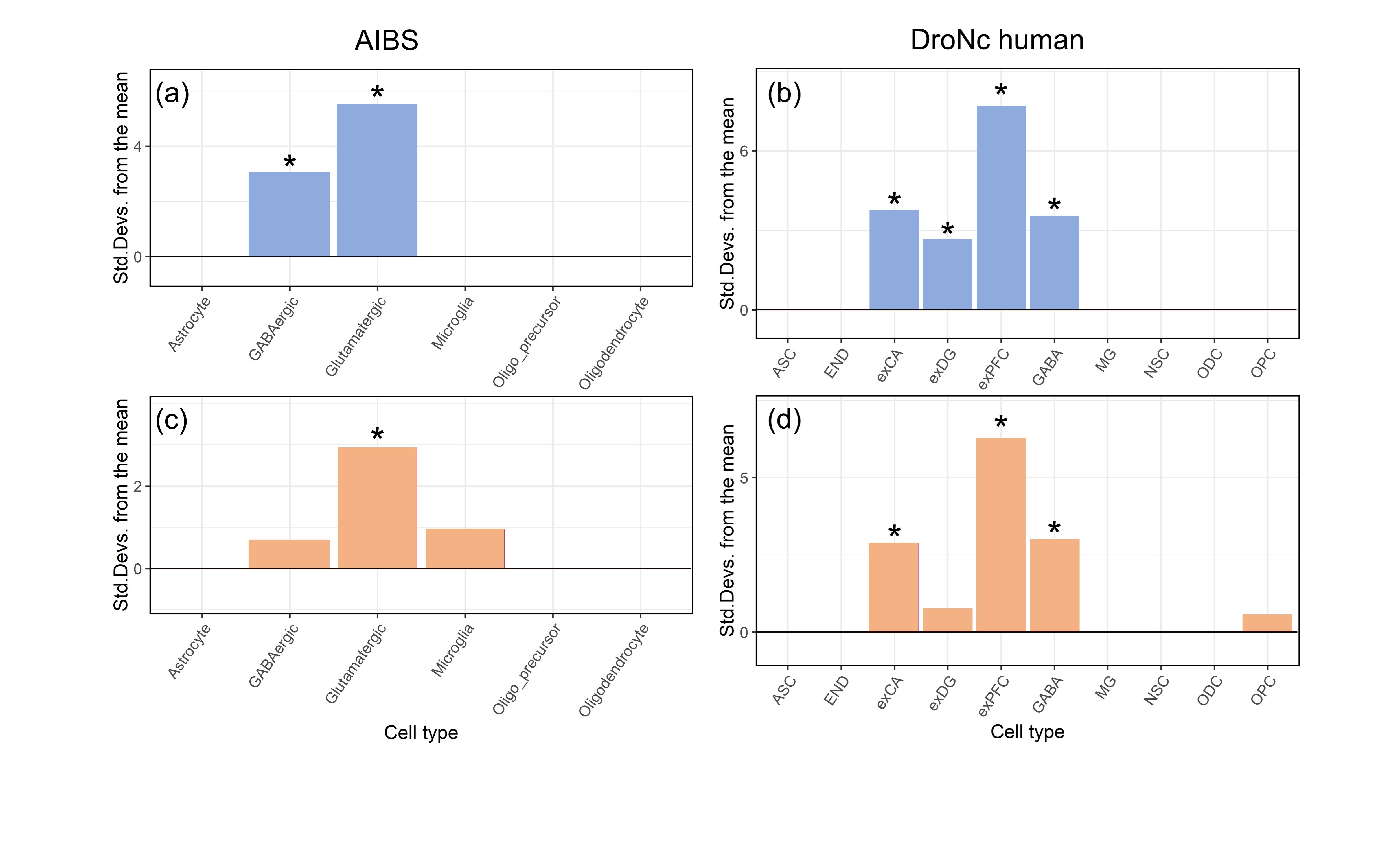
We found that upweighted genes of χdia and χpara both showed significantly greater expression in exCA, exPFC and glutamatergic neurons (Fig. 3). The finding of relative enrichment of exCA in regions with high levels of Aβ and iron is intriguing, as hippocampal CA1 pyramidal projection neurons play a key role in the onset of cognitive impairment during the early phases of AD22. In addition, AD is generally known to be toxic to glutamatergic circuits23, Aβ-induced toxicity was selective for glutamatergic24.
Conclusion
Regional increases in diamagnetic and paramagnetic susceptibility in AD, most likely due to Aβ aggregation and brain iron accumulation, respectively in hippocampus, frontal, cingulate and occipital cortices were explored using APART-QSM. In these regions, we found higher intrinsic levels of gene expression relating to AD associated biological processes and cells. These findings provide valuable insights into the underlying neurodegeneration in AD and the specific vulnerabilities of affected brain regions.Acknowledgements
This work was supported by the National Key R&D Program of China (2020AAA0109502), the National Key R&D Program of China (2021ZD0200500) , National Natural Science Foundation of China (82372036), the Fundamental Research Funds for the Central Universities (226-2023-00095),Open Research Fund of the State Key Laboratory of Cognitive Neuroscience and Learning (CNLZD2001), Key Research Project of Zhejiang Lab (No. 2022ND0AC01).
We thank Lei Zhang, Peiran Jiang from the Core Facilities, Zhejiang University School of Medicine for their technical support.
References
1. Benilova I, Karran E, De Strooper B. The toxic Aβ oligomer and Alzheimer’s disease: An emperor in need of clothes. Nat Neurosci. 2012;15(3):349-357.
2. Becerril-Ortega J, Bordji K, Fréret T, Rush T, Buisson A. Iron overload accelerates neuronal amyloid-β production and cognitive impairment in transgenic mice model of Alzheimer’s disease. Neurobiol Aging. 2014;35(10):2288-2301.
3. Wärmländer SKTS, Österlund N, Wallin C, et al. Metal binding to the amyloid-β peptides in the presence of biomembranes: potential mechanisms of cell toxicity. J Biol Inorg Chem. 2019;24(8):1189-1196.
4. Gong NJ, Dibb R, Bulk M, van der Weerd L, Liu C. Imaging beta amyloid aggregation and iron accumulation in Alzheimer’s disease using quantitative susceptibility mapping MRI. Neuroimage. 2019;191(100):176-185.
5. Ahmed M, Chen J, Arani A, et al. The diamagnetic component map from quantitative susceptibility mapping (QSM) source separation reveals pathological alteration in Alzheimer’s disease-driven neurodegeneration. Neuroimage. 2023;280(January):120357.
6. Zhao Z, Zhang L, Wen Q, et al. The effect of beta-amyloid and tau protein aggregations on magnetic susceptibility of anterior hippocampal laminae in Alzheimer’s diseases. Neuroimage. 2021;244(July):118584.
7. Li Z, Feng R, Liu Q, et al. APART-QSM: An improved sub-voxel quantitative susceptibility mapping for susceptibility source separation using an iterative data fitting method. Neuroimage. 2023;274:120148.
8. Yao J, Tendler BC, Zhou Z, et al. Both noise‐floor and tissue compartment difference in diffusivity contribute to FA dependence on b‐value in diffusion MRI. Hum Brain Mapp. 2023;44(4):1371-1388.
9. Glasser MF, Coalson TS, Robinson EC, et al. A multi-modal parcellation of human cerebral cortex. Nature. 2016;536(7615):171-178.
10. Avants BB, Tustison N, Song G. Advanced Normalization Tools ( ANTS ). 2009:1-29.
11. Thomas GEC, Zarkali A, Ryten M, et al. Regional brain iron and gene expression provide insights into neurodegeneration in Parkinson’s disease. Brain. 2021;144(6):1787-1798.
12. Hawrylycz MJ, Lein ES, Guillozet-Bongaarts AL, et al. An anatomically comprehensive atlas of the adult human brain transcriptome. Nature. 2012;489(7416):391-399.
13. Markello RD, Arnatkeviciute A, Poline J-B, Fulcher BD, Fornito A, Misic B. Standardizing workflows in imaging transcriptomics with the abagen toolbox. Elife. 2021;10.
14. Raudvere U, Kolberg L, Kuzmin I, et al. G:Profiler: A web server for functional enrichment analysis and conversions of gene lists (2019 update). Nucleic Acids Res. 2019;47(W1):W191-W198.
15. Skene NG, Grant SGN. Identification of vulnerable cell types in major brain disorders using single cell transcriptomes and expression weighted cell type enrichment. Front Neurosci. 2016;10(JAN):1-11.
16. Carr, R. M., Oranu, A., & Khungar V, Pengse Po1, Erin Delaney1, Howard Gamper2, Miklos Szanti-Kis3, Lee Speight3, LiWei Tu1, Andrey Kosolapov1, E. James Petersson3, Ya-Ming Hou2 and CD. Canonical Genetic Signatures of the Adult Human Brain Michael. Physiol Behav. 2016;176(1):139-148.
17. Habib N, Avraham-Davidi I, Basu A, et al. Massively parallel single-nucleus RNA-seq with DroNc-seq. Nat Methods. 2017;14(10):955-958.
18. Qiu W, Zhang H, Bao A, Zhu K, Huang Y. Standardized Operational Protocol for Human Brain Banking in China. Neurosci Bull. 2019;35(2):270-276.
19. Van Bergen JMG, Li X, Quevenco FC, et al. Simultaneous quantitative susceptibility mapping and Flutemetamol-PET suggests local correlation of iron and β-amyloid as an indicator of cognitive performance at high age. Neuroimage. 2018;174(February):308-316.
20. Butterfield DA, Reed T, Newman SF, Sultana R. Roles of amyloid β-peptide-associated oxidative stress and brain protein modifications in the pathogenesis of Alzheimer’s disease and mild cognitive impairment. Free Radic Biol Med. 2007;43(5):658-677.
21. Hagemeier J, Geurts JJG, Zivadinov R. Brain iron accumulation in aging and neurodegenerative disorders. Expert Rev Neurother. 2012;12(12):1467-1480.
22. Counts SE, Alldred MJ, Che S, Ginsberg SD, Mufson EJ. Synaptic gene dysregulation within hippocampal CA1 pyramidal neurons in mild cognitive impairment. Neuropharmacology. 2014;79:172-179.
23. Greenamyre JT, Maragos WF, Albin RL, Penney JB, Young AB. Glutamate transmission and toxicity in alzheimer’s disease. Prog Neuropsychopharmacol Biol Psychiatry. 1988;12(4):421-430.
24. Vazin T, Ball KA, Lu H, et al. Efficient derivation of cortical glutamatergic neurons from human pluripotent stem cells: A model system to study neurotoxicity in Alzheimer’s disease. Neurobiol Dis. 2014;62:62-72.
Figures