3889
Brain maps of pCO2, pO2 and pH in aging via homeostatic modeling of neuroimaging data across the lifespan1Radiology, Center for Magnetic Resonance Research (CMRR), University of Minnesota, Minneapolis, MN, United States, 2Netabolics, Rome, Italy, 3Neurology, University of Arkansas, Little Rock, AZ, United States, 4Cell Biology and Physiology, University of New Mexico, Albuquerque, NM, United States, 5Radiology, Magnetic Resonance Research Center (MRRC), Yale University, New Haven, CT, United States, 6Psychiatry, Yale University, New Haven, CT, United States, 7Biomedical Engineering, Yale University, New Haven, CT, United States, 8Anesthesiology, Yale University, New Haven, CT, United States, 9Centro Ricerche Enrico Fermi, Rome, Italy, 10Fondazione Santa Lucia IRCCS, Rome, Italy, 11Psychology, Vanderbilt University, Nashville, TN, United States, 12Department of Neurology, First Faculty of Medicine, Charles University and General University Hospital, Prague, Czech Republic
Synopsis
Keywords: Aging, Aging, Brain, modelling, metabolism, arterial spin labeling, neurovascular coupling, oxygen extraction fraction
Motivation: How loss of vascular health in aging leads to loss of brain function remains unexplained, since paradoxically brain energy consumption is close to normal.
Goal(s): Our goal is to determine whether the loss of vascular health impacts the tissue accumulation of metabolic nutrients and waste products, which may interfere with brain function.
Approach: We exploited Homeostatic Modeling to obtain brain maps of pCO2, pO2 and pH from two retrospective PET and MRI datasets.
Results: Our results show that reduced vascular health in the elderly leads to regional loss of pCO2, pO2, and pH homeostasis of potential clinical significance.
Impact: Determining whether impaired vascular health results into loss of homeostasis of metabolic waste products is critical to guide interventions that improve or preserve brain health in aging and beyond, including neurological conditions such as dementia, stroke, and traumatic brain injury.
Introduction
Altered cerebrovascular health [1-4] and brain metabolism [5, 6] strongly correlate with the aging process. Reduced regional cerebral blood flow (CBF) correlates with cognitive function decline in the elderly [3] independently of structural changes [7]. Yet, despite the reduced CBF, there is adequate energy supply, as resting awake oxygen consumption rate (CMRO2) decreases minimally with age [5, 8]. The mechanisms of action by which loss of vascular health in aging leads to loss of brain function thus remain unexplained. CBF is required not only to deliver nutrients, but also to clear the byproducts of energy metabolism, namely CO2 from glucose oxidation and protons produced by glycolysis of glucose and glycogen to lactate. Notably, studies in humans and preclinical models have shown that brain electrical activity is impacted by acute less than 4% (2 torr) changes in blood pCO2, and 0.01 shifts in pH, and changes can even be detected due to the minute pCO2 variations during the respiration cycle [9, 10]. Small changes in pO2 can also impact brain electrical activity and cognitive performance [11]. Based on this premise, our goal was thus to determine whether the loss of vascular health in aging can lead to functionally significant loss of homeostasis of pCO2, pO2 and pH which may interfere with brain function. Towards achieving this goal, we exploited a recently introduced theoretical framework [12], here named Homeostatic Modeling (HoMod). The core component of HoMod for calculating capillary and venous pO2, pCO2, and pH was previously validated from whole cerebrum arteriovenous measurements [10].Methods
HoMod models the increase in proton and lactate production due to neurometabolic coupling, and demonstrates that if it is not compensated for by a higher rate of clearance by CBF, it leads to a decrease in blood and tissue pH and a resultant increase in pCO2 via the carbonic anhydrase reaction [10]. The model further includes the role of red blood cells in regulating vascular, and consequently tissue, pCO2 and pH as well as pO2. HoMod was here extended to enable regional mapping of pO2, pCO2, and pH based on 3D neuroimaging data, and was then applied to two retrospective datasets across the lifespan, one with PET measurements of Oxygen Extraction Fraction (OEF) from CBF and CMRO2 [5], and one with MRI measurements of CBF obtained with Pseudo-Continuous Arterial Spin Labeling [13].Results
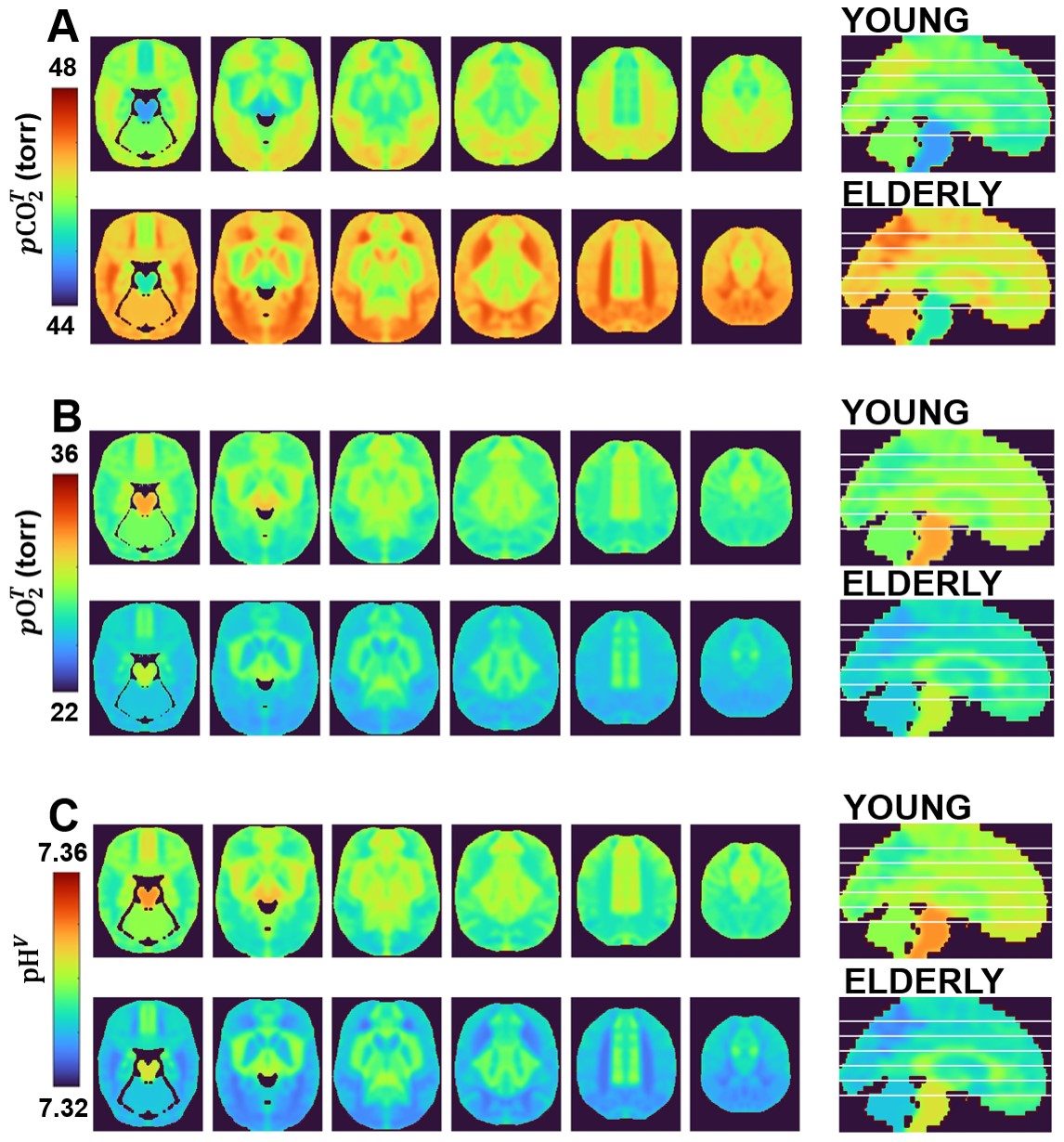
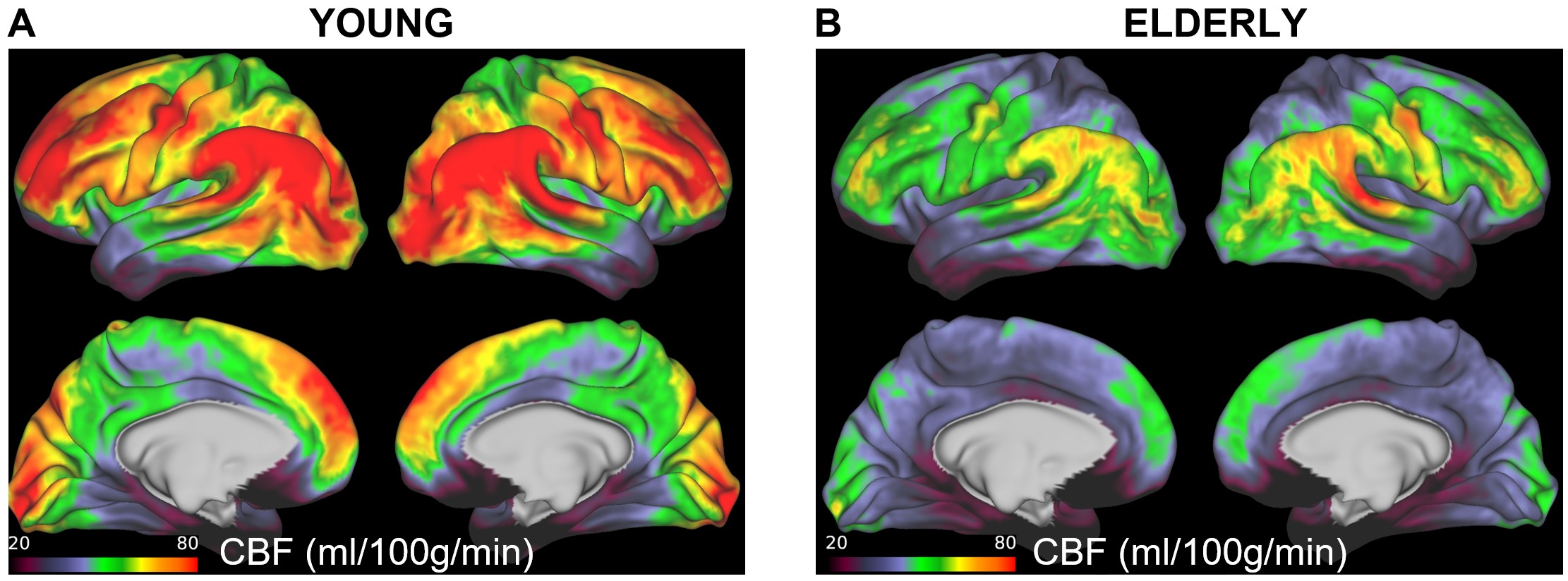
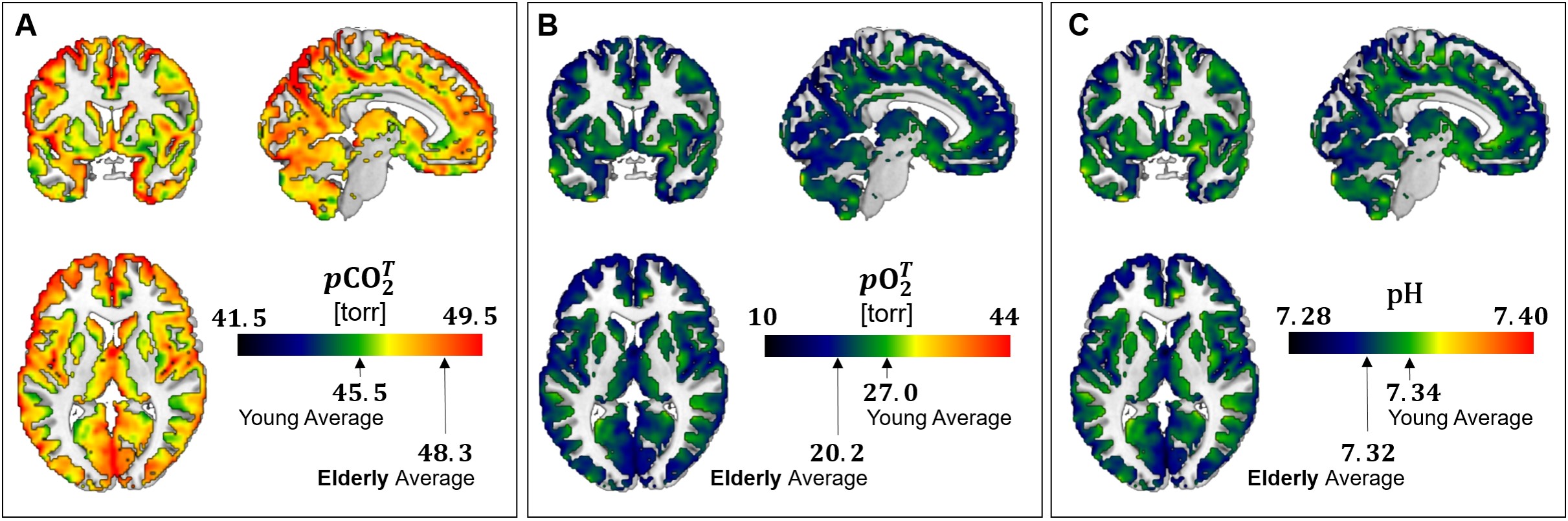
Figure 1 shows regional maps of pCO2, pO2, and pH calculated using HoMod to analyze the PET results reported by Goyal et al. [5] of differences in CBF and CMRO2 across ages between an elderly (61-75 years old, N=66) and a young (20-40 years old, N=66) group of participants. Due to the close to constant OEF in the young group, there is little regional variation in pCO2, pO2, and pH. However, in the elderly, due to reduced CBF (and resultant increased OEF) global and regional increases in pCO2 and decreases in pO2 and pH were found. These age-associated changes were large enough to potentially interfere with normal brain activity. The CBF MRI dataset confirmed global and regional CBF decreases in an elderly (65-84 years old, N=22) vs a young (65-84 years old, N=22) group of participants (Figure 2). Also in this case, HoMod calculated comparable loss of homeostasis from CBF-MRI data in elderly (Figure 3) when assuming similar regional CMRO2 in young and elderly subjects as previously established [5, 8].Discussion
Prior to this analysis, the potential loss of pCO2, pO2, and pH homeostasis due to impaired CBF with aging was not appreciated. It has been traditionally believed that CBF ensures excess of nutrients supply over neuronal demand leading to low OEF, but more recent findings have shown that the low OEF is rather due to the brain’s low capillary density limiting oxygen transport. This paradox of excess supply and inefficient extraction was described in an expert review as “neurovascular coupling – motive unknown” [14]. However, based on the findings of this study, we advance that the main motive of neurovascular coupling is ensuring homeostasis of pCO2, pO2 and pH. If vascular health is impaired, as it occurs in aging, an abnormal accumulation of waste products may be a significant factor in loss of brain function.Conclusion
We conclude that homeostatic modeling enables calculating brain maps of pCO2, pO2 and pH from routine neuroimaging data. Such an opportunity is critical for appreciating mechanisms of action linking loss of vascular health with loss of brain activity in aging, as investigated in this study, and beyond.Acknowledgements
NIH grants U01AG052564, R01AG055591, R01MH109159, R01NS087568, R01NS100106, P41EB027061, 1S10OD017974.References
1. Zimmerman B, Rypma B, Gratton G, et al. Age-related changes in cerebrovascular health and their effects on neural function and cognition: A comprehensive review. Psychophysiology 2021; 58: e13796.
2. Hutchison JL, Lu H and Rypma B. Neural mechanisms of age-related slowing: the DeltaCBF/DeltaCMRO2 ratio mediates age-differences in BOLD signal and human performance. Cereb Cortex 2013; 23: 2337-2346.
3. Wu S, Tyler LK, Henson RNA, et al. Cerebral blood flow predicts multiple demand network activity and fluid intelligence across the adult lifespan. Neurobiol Aging 2023; 121: 1-14.
4. Kim D, Hughes TM, Lipford ME, et al. Relationship Between Cerebrovascular Reactivity and Cognition Among People With Risk of Cognitive Decline. Front Physiol 2021; 12: 645342.
5. Goyal MS, Vlassenko AG, Blazey TM, et al. Loss of Brain Aerobic Glycolysis in Normal Human Aging. Cell Metab 2017; 26: 353-360 e353.
6. Uthayakumar B, Soliman H, Bragagnolo ND, et al. Age-associated change in pyruvate metabolism investigated with hyperpolarized (13) C-MRI of the human brain. Hum Brain Mapp 2023; 44: 4052-4063.
7. Chen JJ, Rosas HD and Salat DH. Age-associated reductions in cerebral blood flow are independent from regional atrophy. Neuroimage 2011; 55: 468-478.
8. Aanerud J, Borghammer P, Chakravarty MM, et al. Brain energy metabolism and blood flow differences in healthy aging. J Cereb Blood Flow Metab 2012; 32: 1177-1187.
9. Bullock T, Giesbrecht B, Beaudin AE, et al. Effects of changes in end-tidal PO(2) and PCO(2) on neural responses during rest and sustained attention. Physiol Rep 2021; 9: e15106.
10. Driver ID, Whittaker JR, Bright MG, et al. Arterial CO2 Fluctuations Modulate Neuronal Rhythmicity: Implications for MEG and fMRI Studies of Resting-State Networks. J Neurosci 2016; 36: 8541-8550.
11. Shaw DM, Cabre G and Gant N. Hypoxic Hypoxia and Brain Function in Military Aviation: Basic Physiology and Applied Perspectives. Front Physiol 2021; 12: 665821.
12. DiNuzzo M, Dienel GA, Behar KL, et al. Neurovascular coupling is optimized to compensate for the increase in proton production from nonoxidative glycolysis and glycogenolysis during brain activation and maintain homeostasis of pH, pCO2 and pO2. J Neurochem 2023 May 7:10.1111/jnc.15839.
13. Harms MP, Somerville LH, Ances BM, et al. Extending the Human Connectome Project across ages: Imaging protocols for the Lifespan Development and Aging projects. Neuroimage 2018; 183: 972-984.
14. Drew
PJ. Neurovascular coupling: motive unknown. Trends
Neurosci 2022; 45: 809-819.
Figures