3875
How much, and in which directions, do subjects move during MRI scans?1Vanderbilt University Medical Center, Nahville, TN, United States, 2Vanderbilt University, Nashville, TN, United States, 3Vanderbilt University Medical Center, Nashville, TN, United States, 4Vanderbilt Memory and Alzheimer's Center, Nashville, TN, United States, 5Laboratory of Behavioral Neuroscience, National Institute on Aging, Baltimore, MD, United States, 6The BIOCARD Study Team, Baltimore, MD, United States, 7The Alzheimer’s Disease Neuroimaging Initiative (ADNI), Los Angeles, CA, United States, 8University of Southern California, Los Angeles, CA, United States
Synopsis
Keywords: Aging, Brain, Motion, Artifacts, Diffusion
Motivation: Subject motion can cause artifacts in MR images and biases in subsequent quantification.
Goal(s): A thorough characterization of motion across multiple cohorts/consortiums has not been performed, particularly with diffusion MRI data.
Approach: We use diffusion MRI data from 14,440 subjects from 10 consortiums (aged 0-100) to characterize (1) how much subjects move during scanning; (2) what kind of head motion is most common; (3) whether motion increases as acquisition proceeds; (4) if motion is associated with age; or (5) with cognitive decline.
Results: Knowledge of the magnitude and direction of motion provides guidance for motion mitigation strategies and informed preprocessing strategies.
Impact: Subject motion induces artifacts and biases in MR images. Characterizing motion magnitude and directions, as well as motion correlates, informs and improves motion mitigation strategies and image processing pipelines.
Introduction
It is well-known that subject motion induces artifacts in MR images that have a downstream impact on image-derived metrics. In structural studies (T1/T2-weighted), motion results in misestimates geometry [1, 2]. In functional studies, motion causes image artifacts that confounds analysis [3] and introduces spurious network correlations [4]. In diffusion studies, motion introduces misalignment between volumes, signal dropout, and erroneous signal attenuation [5] which biases measures and can artifactually create group differences [6]. While it is well-known that different magnitudes, directions, and frequencies of motion can cause different artifacts, a thorough characterization of motion across a population has not been performed, particularly with diffusion MRI data.Methods
We used 10 large consortium datasets to study subject motion across a population (0-100 years). We chose diffusion MRI data which involved collection of multiple volumes over several minutes and allowed characterizing motion: ADNI (N=2521) [7], BIOCARD (N=974) [8], ROSMAP (N=1973) [9], BLSA (N=5424) [10], ICBM (N=140) [11], OASIS3 (N=1794) [12], OASIS4 (N=603) [13], ABVIB (N=63) , CAMCAN (N=305) [14], BabyHCP (N=494) [15]. All datasets had varying scan parameters (TR/TE/Number of Volumes/Resolution), different scan durations, and possibly different sensitivity to motion. Preprocessing was performed to attenuate susceptibility, motion, and eddy currents. Derived measures of motion included relative mean displacement (mm), translation in Left/Right, Posterior/Anterior, Interior/Superior directions (mm), and rotation around L/R, P/A, I/S axis (radians). All measures were derived per imaging volume, and also normalized to scan duration (i.e., mean displacement per minute).Results
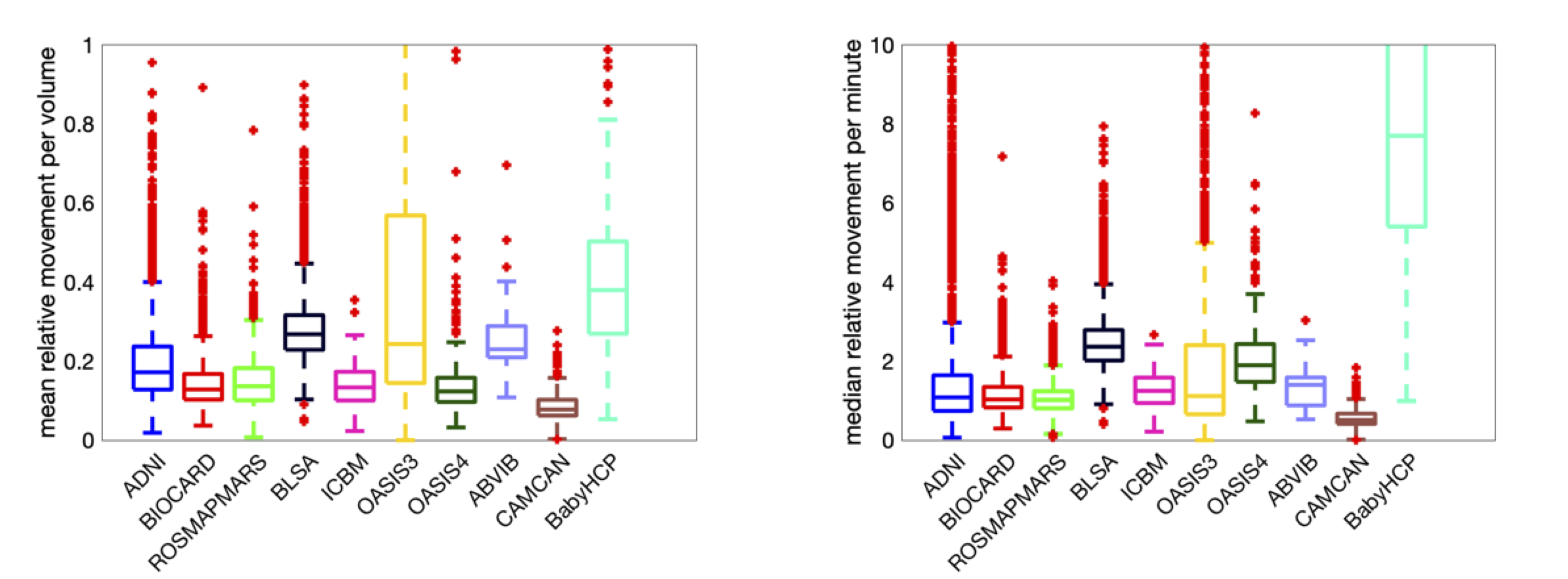
How much do subjects move during scans?Figure 1 shows the mean relative movement per volume and mean relative movement per minute for all datasets. On average, voxels in the brain move ~0.1-0.2mm/volume or ~1-2mm/minute, with differences observable across datasets, and high movement for the BabyHCP cohort. Additional analysis shows the mean movement is greater than the median movement, which suggest frequent smaller movements are more common than occasional large movements.
What kind of head motion is most common during scans?
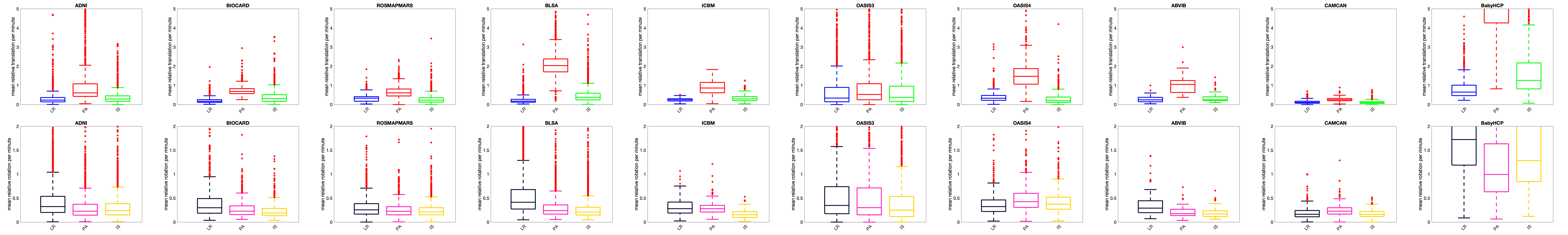
Figure 2 shows measures of brain translation/rotation for each dataset. Translation tends to be greatest in the AP direction, whereas rotation is largest around the LR axis (i.e., head nodding), for most datasets.
Does motion get worse as acquisition proceeds?
The cumulative motion over time for all subjects in each dataset is plotted in Figure 3. In most cases, motion largely linearly increases over the full scan duration.
Is motion related to age?
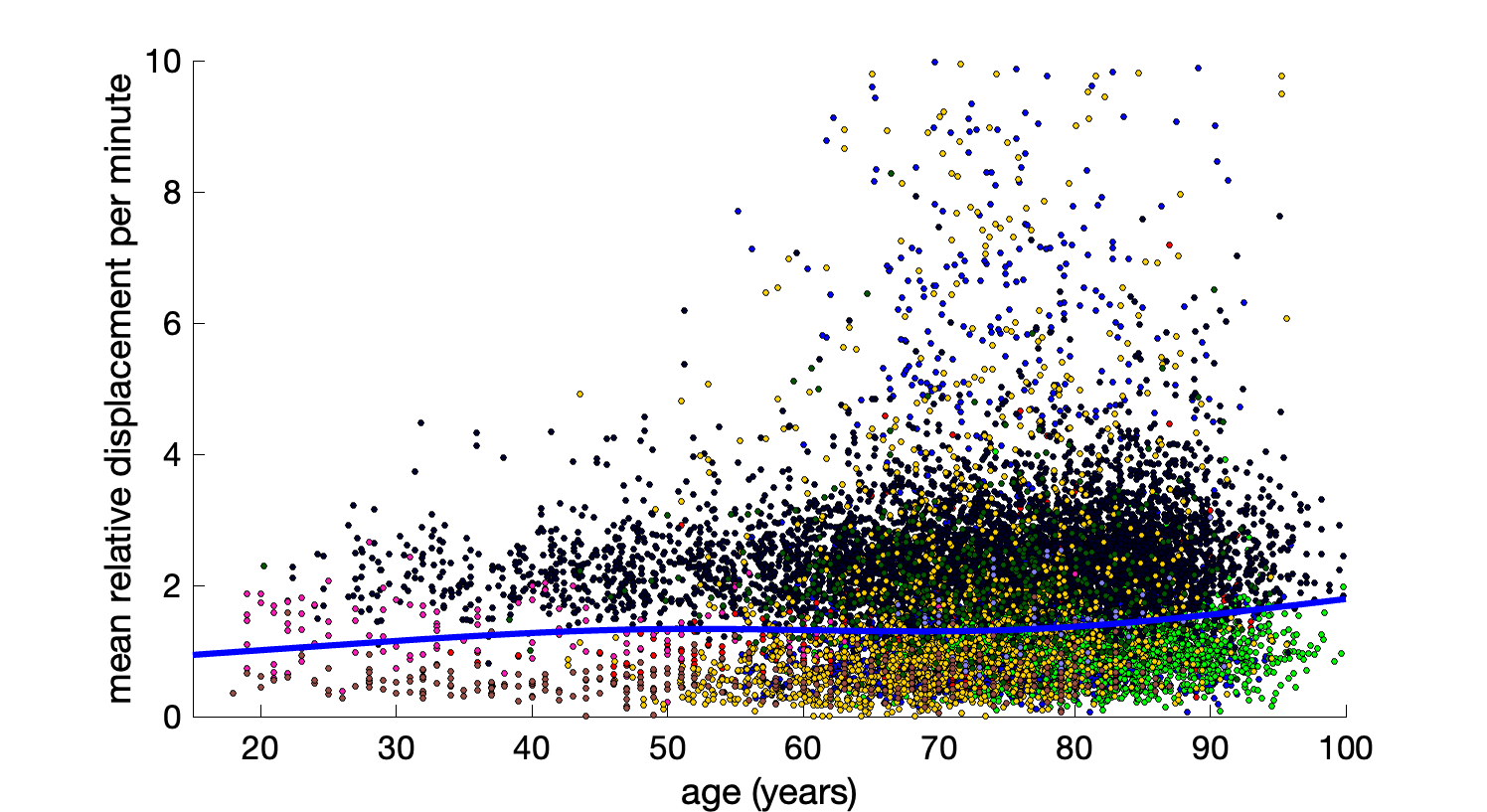
A plot of mean displacement against age for all subjects is shown in Figure 4, with data fit to a covariate-adjusted restricted cubic spline regression. Motion tends to increase with age, and the number of outliers with large motion also increases with age. Note that BabyHCP is not shown in this plot as it has the largest motion.
Does cognitive decline contribute to motion?
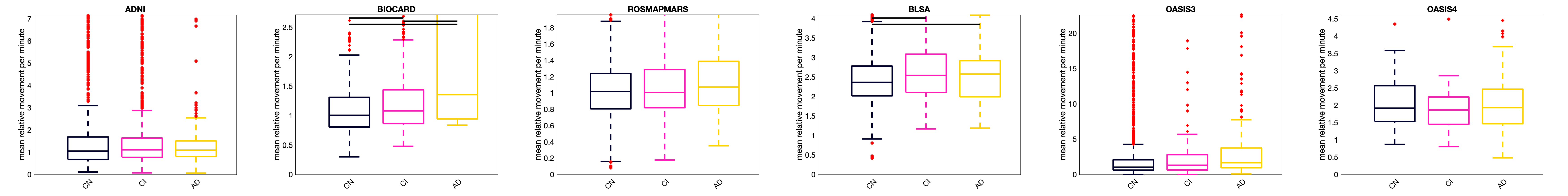
Figure 5 shows motion for cognitively normal, mild cognitive impairmen), and Alzheimer’s subjects for each dataset. Motion tends to increase with increasing cognitive impairment, with differences in the BIOCARD and BLSA datasets showing statistically significant increases.
Discussion
We characterize motion across several large cohorts and consortiums, across a large age range and find that on average, volunteers in the scanner move ~1-2mm per minute, with frequent smaller movements more common than occasional large movements. There are also meaningful site differences – meaning that in addition to scanner effects, choices in subject positioning and immobilization strategies, as well as scan protocol, affect the observed motion. Further analysis should investigate how choices in scan length, number of volumes, and time between scans (TR) influence observed motion and subsequent artifacts.Translation occurs most in the AP direction, while rotation occurs round the LR axis. These are possibly the least constrained directions of motion, as earpads/constraints prevent LR translation and SI rotation. While intuitive, this observation has not been well described in literature, and we show that these results generalize across cohorts/scan sites.
Qualitatively, motion does not seem to increase as the scan proceeds, although further analysis into the trends over scan duration are needed.
Motion increases with age. Moreover, outliers become more frequent above the age of 60. This re-emphasizes the fact that motion can cause spurious, non-biological, differences in cohorts if not properly accounted for. Similar results are observed in those with cognitive impairment, with motion increasing along the impairment spectrum.
We expect these results to generalize to all data/contrasts, as there is little fundamental difference to the subject between diffusion and functional MRI. Future studies should confirm similar results in functional imaging, and investigate effects of motion on microstructure and connectivity estimates derived from diffusion MRI.
Acknowledgements
This work was supported by the National Science Foundation Career Award #1452485, the National Institutes of Health under award numbers R01EB017230, K01EB032898, S10OD021771 01, U54DK134302, U54EY032442, and in part by ViSE/VICTR VR3029 and the National Center for Research Resources, Grant UL1 RR024975–01. The BLSA is supported by the Intramural Research Program, National Institute on Aging, NIH. The BIOCARD study is supported by a grant from the National Institute on Aging (NIA): U19-AG03365. The BIOCARD Study consists of 7 Cores and 2 projects with the following members: (1) The Administrative Core (Marilyn Albert, Corinne Pettigrew, Barbara Rodzon); (2) the Clinical Core (Marilyn Albert, Anja Soldan, Rebecca Gottesman, Corinne Pettigrew, Leonie Farrington, Maura Grega, Gay Rudow, Rostislav Brichko, Scott Rudow, Jules Giles, Ned Sacktor); (3) the Imaging Core (Michael Miller, Susumu Mori, Anthony Kolasny, Hanzhang Lu, Kenichi Oishi, Tilak Ratnanather, Peter vanZijl, Laurent Younes); (4) the Biospecimen Core (Abhay Moghekar, Jacqueline Darrow, Alexandria Lewis, Richard O’Brien); (5) the Informatics Core (Roberta Scherer, Ann Ervin, David Shade, Jennifer Jones, Hamadou Coulibaly, Kathy Moser, Courtney Potter); the (6) Biostatistics Core (Mei-Cheng Wang, Yuxin Zhu, Jiangxia Wang); (7) the Neuropathology Core (Juan Troncoso, David Nauen, Olga Pletnikova, Karen Fisher); (8) Project 1 (Paul Worley, Jeremy Walston, Mei-Fang Xiao), and (9) Project 2 (Mei-Cheng Wang, Yifei Sun, Yanxun Xu. Data collection and sharing for ADNI were supported by National Institutes of Health Grant U01-AG024904 and Department of Defense (award number W81XWH-12-2-0012). ADNI is also funded by the National Institute on Aging, the National Institute of Biomedical Imaging and Bioengineering, and through generous contributions from the following: AbbVie, Alzheimer’s Association; Alzheimer’s Drug Discovery Foundation; Araclon Biotech; BioClinica, Inc.; Biogen; Bristol-Myers Squibb Company; CereSpir, Inc.; Cogstate; Eisai Inc.; Elan Pharmaceuticals, Inc.; Eli Lilly and Company; EuroImmun; F. Hoffmann-La Roche Ltd and its affiliated company Genentech, Inc.; Fujirebio; GE Healthcare; IXICO Ltd.; Janssen Alzheimer Immunotherapy Research & Development, LLC.; Johnson & Johnson Pharmaceutical Research & Development LLC.; Lumosity; Lundbeck; Merck & Co., Inc.; Meso Scale Diagnostics, LLC.; NeuroRx Research; Neurotrack Technologies; Novartis Pharmaceuticals Corporation; Pfizer Inc.; Piramal Imaging; Servier; Takeda Pharmaceutical Company; and Transition Therapeutics. The Canadian Institutes of Health Research is providing funds to support ADNI clinical sites in Canada. Private sector contributions are facilitated by the Foundation for the National Institutes of Health (www.fnih.org). The grantee organization is the Northern California Institute for Research and Education, and the study is coordinated by the Alzheimer’s Therapeutic Research Institute at the University of Southern California. ADNI data are disseminated by the Laboratory for Neuro Imaging at the University of Southern California. Data were provided in part by OASIS for the OASIS-3 cohort: Longitudinal Multimodal Neuroimaging: Principal Investigators: T. Benzinger, D. Marcus, J. Morris; NIH P30 AG066444, P50 AG00561, P30 NS09857781, P01 AG026276, P01 AG003991, R01 AG043434, UL1 TR000448, R01 EB009352. AV-45 doses were provided by Avid Radiopharmaceuticals, a wholly owned subsidiary of Eli Lilly. HCP: Data were provided [in part] by the Human Connectome Project, WU- Minn Consortium (Principal Investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at Washington University. The content is solely the responsibility of the authors and does not necessarily represent the official views of the NIH. Data used in preparation of this article were obtained from the Aging Brain: Vasculature, Ischemia, and Behavior Study (ABVIB). As such, the key investigators within the ABVIB contributed to the design and implementation of ABVIB and/or provided data but did not participate in analysis or writing of this report: Helena C. Chui M.D. (Principal Investigator), Charles C. DeCarli, M.D., William G. Ellis, M.D., William J. Jagust, M.D., Joel H. Kramer, Ph.D., Meng Law, M.D., Dan Mungas Ph,D. Bruce R. Reed, Ph.D., Nerses Sanossian, M.D., Michael W. Weiner, M.D. Wendy J. Mack, Ph.D., Harry V. Vinters, M.D., Chris Zarow, Ph.D., Ling Zheng, Ph.D.References
1. Blumenthal, J.D., A. Zijdenbos, E. Molloy, and J.N. Giedd, Motion artifact in magnetic resonance imaging: implications for automated analysis. Neuroimage, 2002. 16(1): p. 89-92.
2. Savalia, N.K., et al., Motion-related artifacts in structural brain images revealed with independent estimates of in-scanner head motion. Hum Brain Mapp, 2017. 38(1): p. 472-492.
3. Friston, K.J., et al., Movement-related effects in fMRI time-series. Magn Reson Med, 1996. 35(3): p. 346-55.
4. Power, J.D., et al., Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. Neuroimage, 2012. 59(3): p. 2142-54.
5. Anderson, A.W. and J.C. Gore, Analysis and correction of motion artifacts in diffusion weighted imaging. Magn Reson Med, 1994. 32(3): p. 379-87.
6. Yendiki, A., et al., Spurious group differences due to head motion in a diffusion MRI study. NeuroImage, 2014. 88: p. 79-90.
7. Jack, C.R., Jr., et al., The Alzheimer's Disease Neuroimaging Initiative (ADNI): MRI methods. J Magn Reson Imaging, 2008. 27(4): p. 685-91.
8. Albert, M., et al., Cognitive changes preceding clinical symptom onset of mild cognitive impairment and relationship to ApoE genotype. Curr Alzheimer Res, 2014. 11(8): p. 773-84.
9. Bennett, D.A., et al., Religious Orders Study and Rush Memory and Aging Project. J Alzheimers Dis, 2018. 64(s1): p. S161-S189.
10. Williams, O.A., et al., Vascular burden and APOE epsilon4 are associated with white matter microstructural decline in cognitively normal older adults. Neuroimage, 2019. 188: p. 572-583.
11. Crawford, K.L., S.C. Neu, and A.W. Toga, The Image and Data Archive at the Laboratory of Neuro Imaging. NeuroImage, 2016. 124: p. 1080-1083.
12. Pamela, J.L., et al., OASIS-3: Longitudinal Neuroimaging, Clinical, and Cognitive Dataset for Normal Aging and Alzheimer Disease. medRxiv, 2019: p. 2019.12.13.19014902.
13. Marcus, D.S., et al., Open Access Series of Imaging Studies (OASIS): cross-sectional MRI data in young, middle aged, nondemented, and demented older adults. J Cogn Neurosci, 2007. 19(9): p. 1498-507.
14. Taylor, J.R., et al., The Cambridge Centre for Ageing and Neuroscience (Cam-CAN) data repository: Structural and functional MRI, MEG, and cognitive data from a cross-sectional adult lifespan sample.Neuroimage, 2017. 144(Pt B): p. 262-269.
15. Howell, B.R., et al., The UNC/UMN Baby Connectome Project (BCP): An overview of the study design and protocol development. Neuroimage, 2019. 185: p. 891-905.
Figures