3479
Cell radius and membrane permeability in the developing brain1Department of Radiology and BRIC, UNC Chapel Hill, Chapel Hill, NC, United States
Synopsis
Keywords: Microstructure, Microstructure, axon radius, membrane permeability, development, postnatal
Motivation: Axon and soma radii and membrane permeability are important features of brain structure and pathology but in-vivo noninvasive measurement of these biomarkers is challenging due to complex tissue architectures and inherent simplifying assumptions in MR biophysical models.
Goal(s): We study membrane permeability and axon and soma radii of the developing brain during the first 5 years of life, the most dynamic and complex postnatal neurodevelopment.
Approach: Different microstructure models were fitted to 389 diffusion MRI scans of 217 subjects. Measurements and developmental trends were compared with histological evidence.
Results: MF-SMSI yields results that are closest to histological expectations with biologically plausible developmental trends.
Impact: Noninvasive measurements of cell physical properties in the developing brain were compared with histological evidence. We present a method that yields results that are closest to histological expectations with biologically plausible developmental trends. The method can potentially facilitate neurodevelopmental studies.
Introduction
Physical properties of neural cells, such as radius and membrane permeability, reflect brain microstructure and pathology. Noninvasive measurement of the properties of complex tissue geometries using MR biophysical models remains challenging. Multiple methods have been proposed to address this challenge but their application to the developing brain, the most dynamic and complex postnatal neurodevelopmental period, is not fully explored. Simplifying assumptions associated with most models could lead to inaccurate measurements. In this work, we study membrane permeability and axon and soma radii of the developing brain during the first 5 years of life. Measurements and developmental trends given by multiple biophysical models were compared with histological evidence.Methods
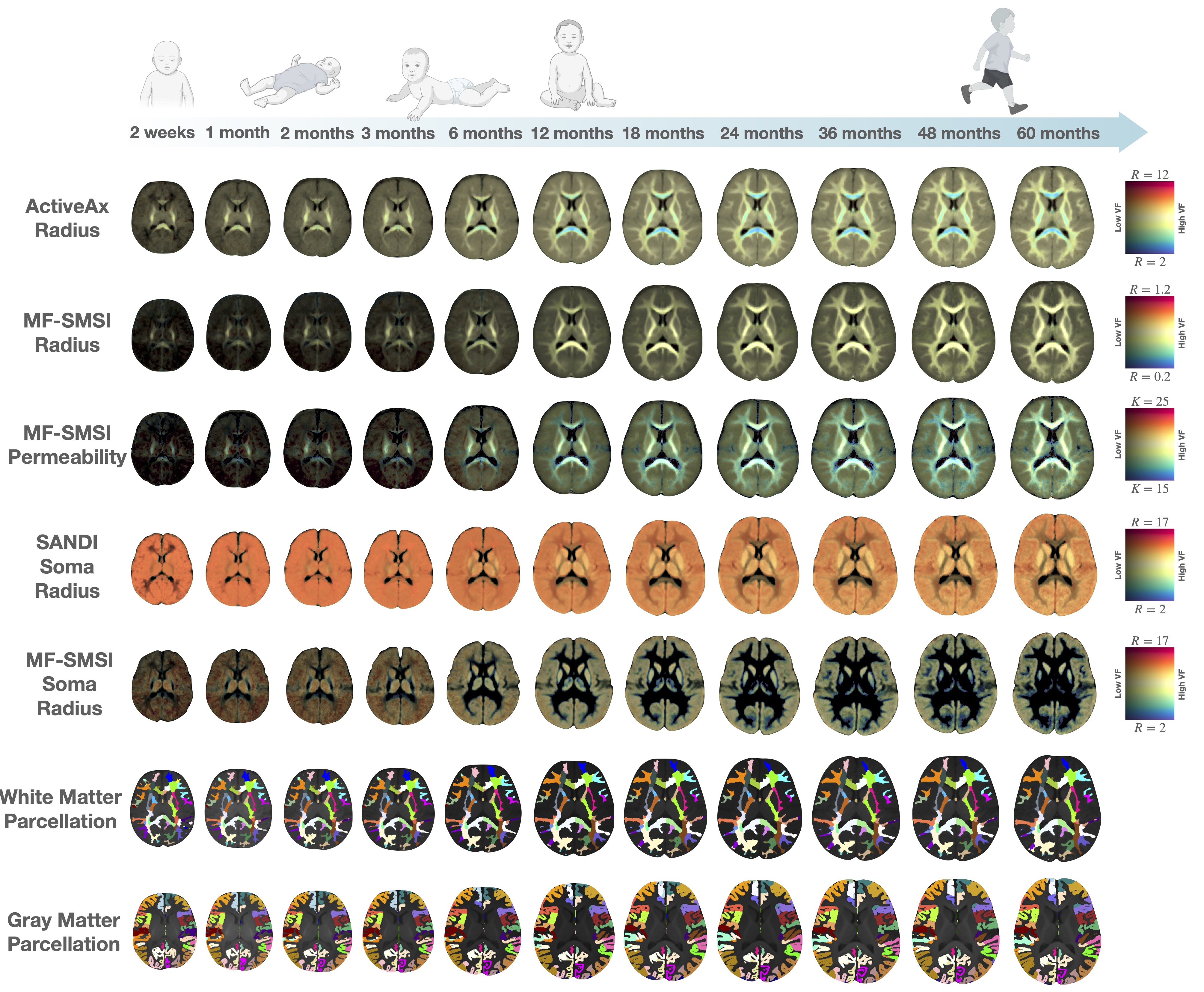
We used a total of 389 6-shell diffusion MRI scans of 217 subjects acquired from birth to 5 years of age using an accelerated longitudinal scheme [4]. Microstructure maps were calculated in native individual space. Age-specific white matter [5] and gray matter [6] parcellations were transferred to the subject native space for regional measurements. The GAMM [7] was fitted to the developmental curves with age as a fixed effect and gender and subject as random effects.We investigated 3 biophysical models:
- ActiveAx [1]: A minimal four-compartment model, including intra-axonal, extra-axonal, CSF, and trapped water compartments, for measuring axonal radius.
- SANDI [2]: A three-compartment model, including intra-neurite, intra-soma, and extra-cellular compartments, for measuring soma radius.
- MF-SMSI [3]: A microstructure fingerprinting model, which represents the signal as a combination of a spectrum of micro-environments, for measuring axon and soma radii as well as axon membrane permeability.
Results
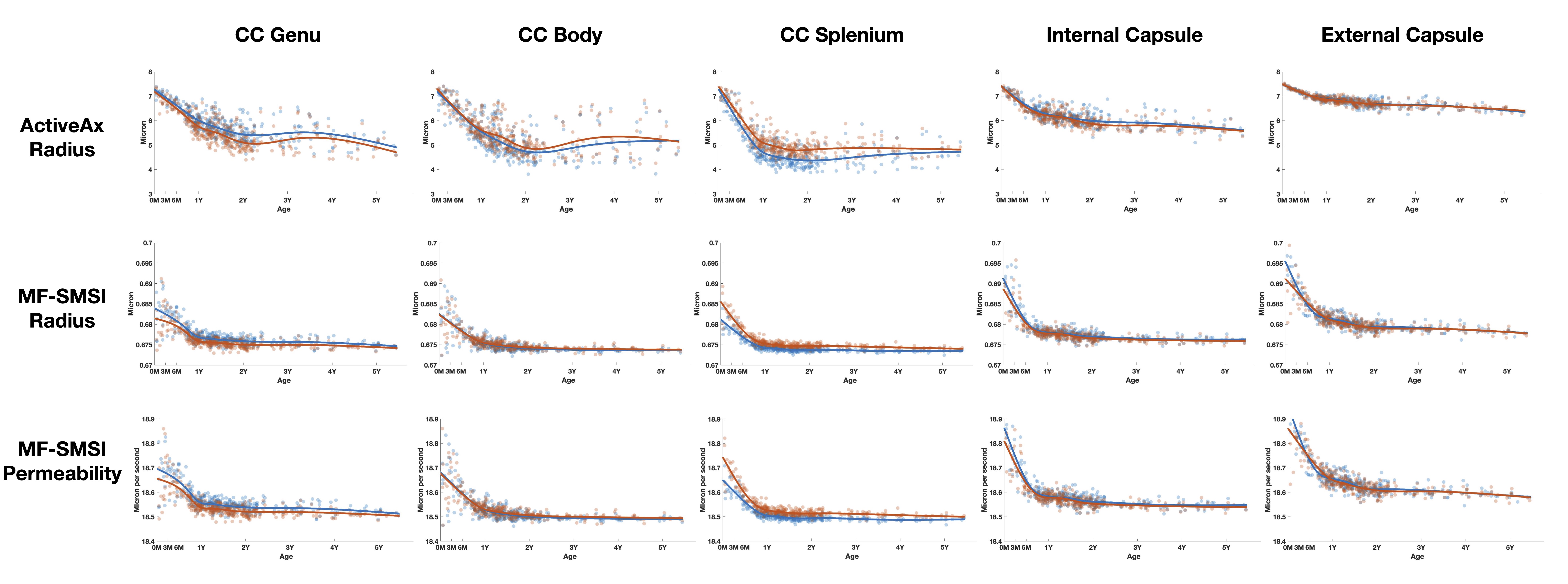
Axon radius:Histological studies [8,9,10] reported a skewed distribution with 95% axons having a radius less than 1 micron. The average human axon radius ranges from 0.59 to 0.8 micron (after accounting for 10-30% shrinkage in ex-vivo samples).
ActiveAx overestimates the axon radius, reporting an average radius of 7 micron at birth and down to 5 micron at 5 years of age (Figs. 1 & 3, 1st row). The axon radius decreases respectively by 14% and 28% (volume decreases 30% and 60%) in the first 6 and 12 months. Biological evidence supporting such drastic reduction in axonal radius remains elusive.
MF-SMSI reports axon radii ranging from 0.67 to 0.69 micron (Figs. 1 & 3,, 2nd row), inline with histological observations. The axon radius decreases by a small margin of 1-2% in the first year and plateaus thereafter.
Both methods show a spatial pattern of radii being larger at the center (e.g., corpus callosum) and smaller at the periphery (e.g., internal and external capsules).
Axon membrane permeability:
MF-SMSI (Figs. 1 & 3, 3rd row) shows 18.5 - 19 um/s membrane permeability, agreeing well with the biological range of 3-30 um/s [8]. Membrane permeability decreases in the first year with greater permeability in peripheral regions than the corpus callosum, in line with temporal and spatial characteristics of myelination in the developing brain.
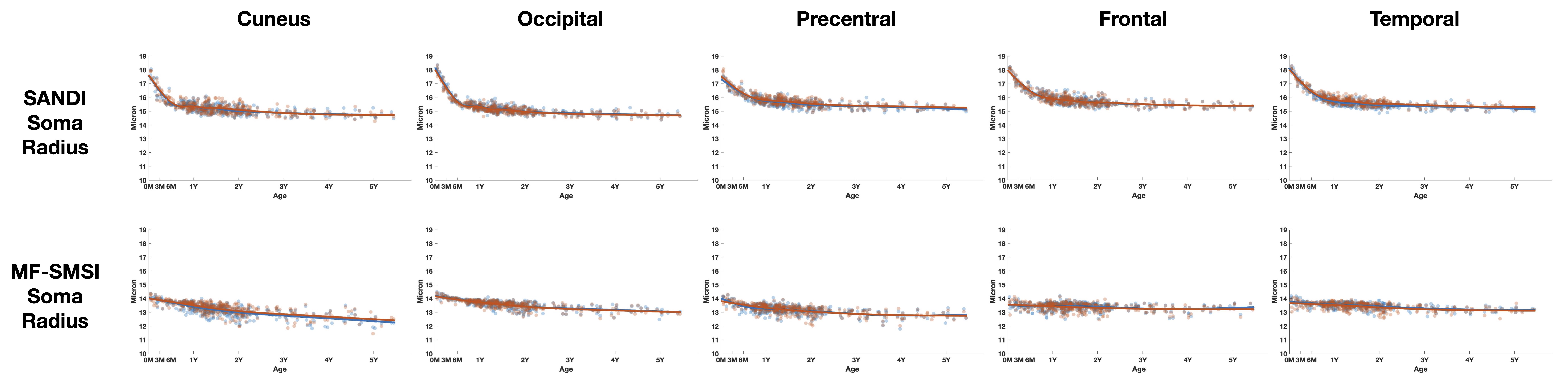
Soma radius:
Human brain cellular morphology database reports an average soma radius of 11 (+- 7) micron [2,11]. Both SANDI and MF-SMSI measurements (Figs. 2 & 3) are within this range with MF-SMSI closer to the ex-vivo measurements (after correction for ex-vivo shrinkage). SANDI estimates an 11% decrease in radius (30% in volume) in the first 6 months, which is challenging to explain for healthy microstructural development. On the contrary, MF-SMSI shows more plausible trends with a marginal decrease of 3% in the first year.
Conclusion
We compared radius measurements and membrane permeability given by different biophysical models for the developing brain with histological data. Overall, MF-SMSI yields results that are closest to histological expectations with biologically plausible developmental trends.Acknowledgements
This work was supported in part by the United States National Institutes of Health (NIH) through grants MH125479 and EB008374.References
1. Alexander DC, Hubbard PL, Hall MG, Moore EA, Ptito M, Parker GJ, Dyrby TB. Orientationally invariant indices of axon diameter and density from diffusion MRI. Neuroimage. 2010 Oct 1;52(4):1374-89.
2. Palombo M, Ianus A, Guerreri M, Nunes D, Alexander DC, Shemesh N, Zhang H. SANDI: a compartment-based model for non-invasive apparent soma and neurite imaging by diffusion MRI. Neuroimage. 2020 Jul 15;215:116835.
3. Huynh KM, Wu Y, Ahmad S, Yap PT. Microstructure Fingerprinting for Heterogeneously Oriented Tissue Microenvironments. InInternational Conference on Medical Image Computing and Computer-Assisted Intervention 2023 Oct 1 (pp. 131-141). Cham: Springer Nature Switzerland.
4. Howell BR, Styner MA, Gao W, Yap PT, Wang L, Baluyot K, Yacoub E, Chen G, Potts T, Salzwedel A, Li G. The UNC/UMN Baby Connectome Project (BCP): An overview of the study design and protocol development. NeuroImage. 2019 Jan 15;185:891-905.
5. Oishi K, Faria A, Jiang H, Li X, Akhter K, Zhang J, Hsu JT, Miller MI, van Zijl PC, Albert M, Lyketsos CG. Atlas-based whole brain white matter analysis using large deformation diffeomorphic metric mapping: application to normal elderly and Alzheimer's disease participants. Neuroimage. 2009 Jun 1;46(2):486-99.
6. Alexander B, Loh WY, Matthews LG, Murray AL, Adamson C, Beare R, Chen J, Kelly CE, Anderson PJ, Doyle LW, Spittle AJ. Desikan-Killiany-Tourville atlas compatible version of M-CRIB neonatal parcellated whole brain atlas: The M-CRIB 2.0. Frontiers in Neuroscience. 2019 Feb 5;13:34.
7. Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM, Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM. GLMM and GAMM. Mixed effects models and extensions in ecology with R. 2009:323-41.
8. Veraart J, Nunes D, Rudrapatna U, Fieremans E, Jones DK, Novikov DS, Shemesh N. Noninvasive quantification of axon radii using diffusion MRI. elife. 2020 Feb 12;9:e49855.
9. Aboitiz F, Scheibel AB, Fisher RS, Zaidel E. Fiber composition of the human corpus callosum. Brain research. 1992 Dec 11;598(1-2):143-53.
10. Caminiti R, Ghaziri H, Galuske R, Hof PR, Innocenti GM. Evolution amplified processing with temporally dispersed slow neuronal connectivity in primates. Proceedings of the National Academy of Sciences. 2009 Nov 17;106(46):19551-6.
11. https://neuromorpho.org
Figures