3471
Axonal fraction imaging on clinical and preclinical dMRI PGSE data1Department of Applied Mathematics and Computer Science, Technical University of Denmark, Kgs. Lyngby, Denmark, 2Danish Research Centre for Magnetic Resonance, Centre for Functional and Diagnostic Imaging and Research, Copenhagen University Hospital Amager and Hvidovre, Copenhagen, Denmark
Synopsis
Keywords: Microstructure, Microstructure, Diffusion
Motivation: We wish to quantify the amount of axons in the brain using clinically-feasible in vivo human diffusion MRI.
Goal(s): We want to estimate the axonal signal fraction using the conventional pulsed gradient spin echo (PGSE) sequence while reducing model degeneracy and minimizing modeling assumptions.
Approach: We model spherical harmonic (SH) coefficients across two high b-value PGSE shells. We calculate ratios between SH l-band power spectra across the shells, relate them analytically to the axonal diffusivities - estimated using machine learning - and with these we calculate the axonal signal fraction.
Results: We report comparable results across preclinical and clinical data and demonstrate methodological feasibility.
Impact: The axonal signal fraction is proportional to the total volume of axons within a voxel and can be used to characterize pathology. This work proposes its estimation with clinically-feasible b-values and with conventional diffusion MRI data while minimizing modeling assumptions.
Introduction
The axonal signal fraction (ASF) is a proxy for brain tissue integrity related to the axonal volume occupancy in every voxel. The ASF estimated from multicompartmental models1 shows good correlation with histology2. However, undesirable modeling assumptions and/or specific acquisition requirements are needed. In particular, when using PGSE data, degeneracy3 may force us to assume a tortuosity model4 and/or fix parameters5. Degeneracy is circumvented using mixtures of multiple/tensor encodings6,7,8 however this may entail mixing together different spectral modulations9, introducing unaccounted heterogeneity in the diffusion weighting. In this work, we use high b-value PGSE data to only model the signal arising from axons10, thus minimizing degeneracy.Axonal diffusion is modeled as a dispersed axisymmetric tensor with axial and radial diffusivity $$$\lambda_{\parallel}$$$ and $$$\lambda_{\perp}$$$. Previously, we showed that at high b-values the powder average signal (commonly used to avoid estimating dispersion) only retains sensitivity to $$$\lambda_{\perp}$$$11. Moreover, since $$$\lambda_{\perp}$$$ is small, we can only estimate it using a preclinical scanner like the Connectome12 because clinically-feasible (in terms of SNR) high b-values do not attenuate the powder average signal sufficiently, leading to the clinical approximation $$$\lambda_{\perp}\approx 0$$$10. We also showed11 that zonal degree (l-band) ratios of SH coefficients (across shells) with $$$l\ge2$$$ retain sensitivity to both $$$\lambda_{\parallel}$$$ and $$$\lambda_{\perp}$$$. These ratios are also rotationally invariant (unaffected by dispersion) and provide additional sensitivity to $$$\lambda_{\perp}$$$. Therefore, in this work we leverage these ratios to break out of the axonal diffusivities estimation conundrum, which allows us to then calculate the ASF.
Methods
We expand each shell at high b-value $$$b_1$$$ and $$$b_2$$$ as13$$ S(b_1, \textbf{n}) = \sum_{l=0, even}^{L}\sum_{m=-l}^{l} c_{lm}(b_1) Y_l^m(\textbf{n})$$
$$ S(b_2, \textbf{n}) = \sum_{l=0, even}^{L}\sum_{m=-l}^{l} c_{lm}(b_2) Y_l^m(\textbf{n})$$
where $$$\textbf{n}$$$ is the gradient direction, $$$Y_l^m(\cdot)$$$ the real spherical harmonics13, and $$$c_{lm}(\cdot)$$$ the coefficients of shells at b-values $$$b_1$$$ and $$$b_2$$$. Using the coefficients and assuming an axonal convolutional model14 we derive the $$$l$$$-band ratios11
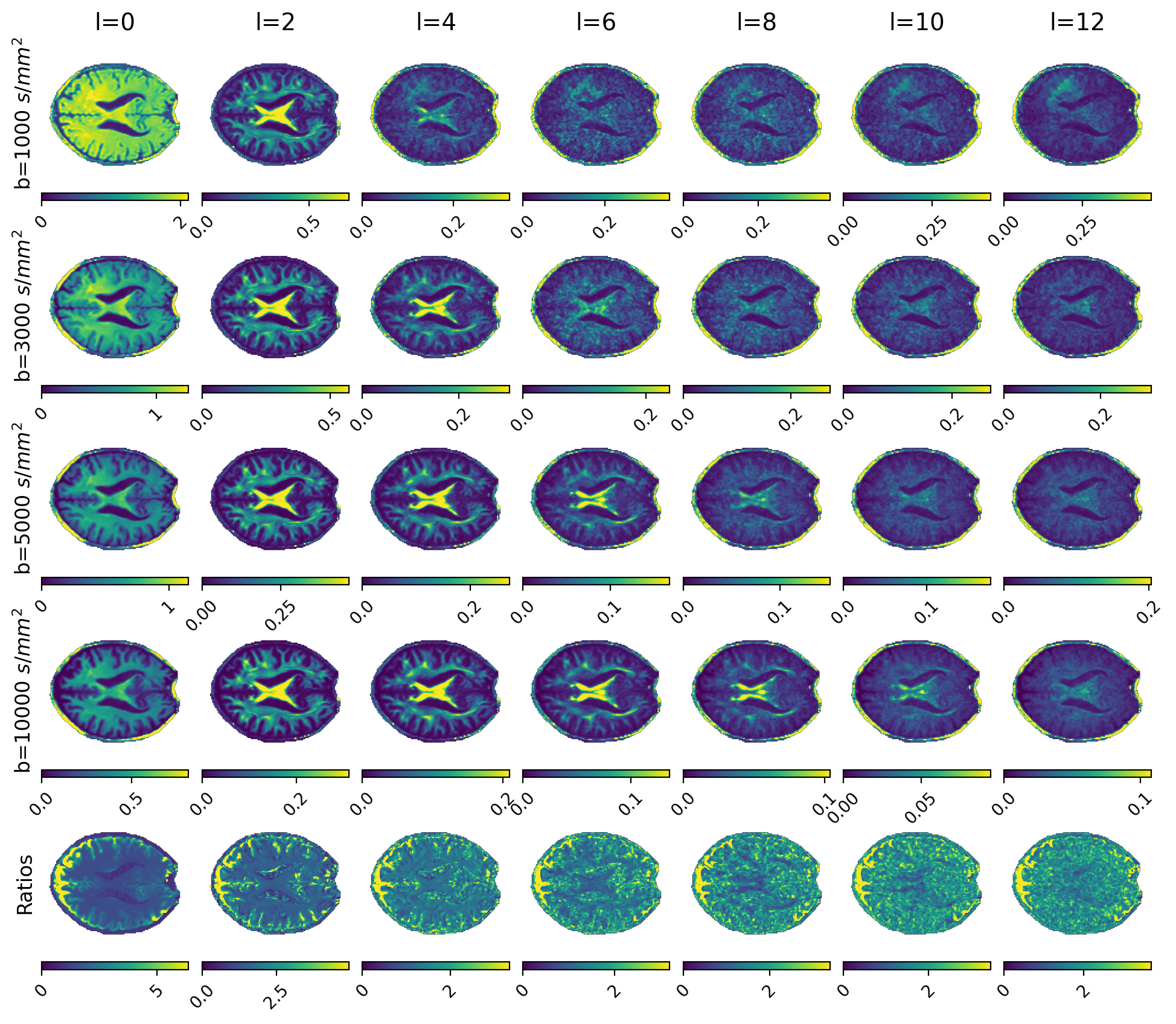
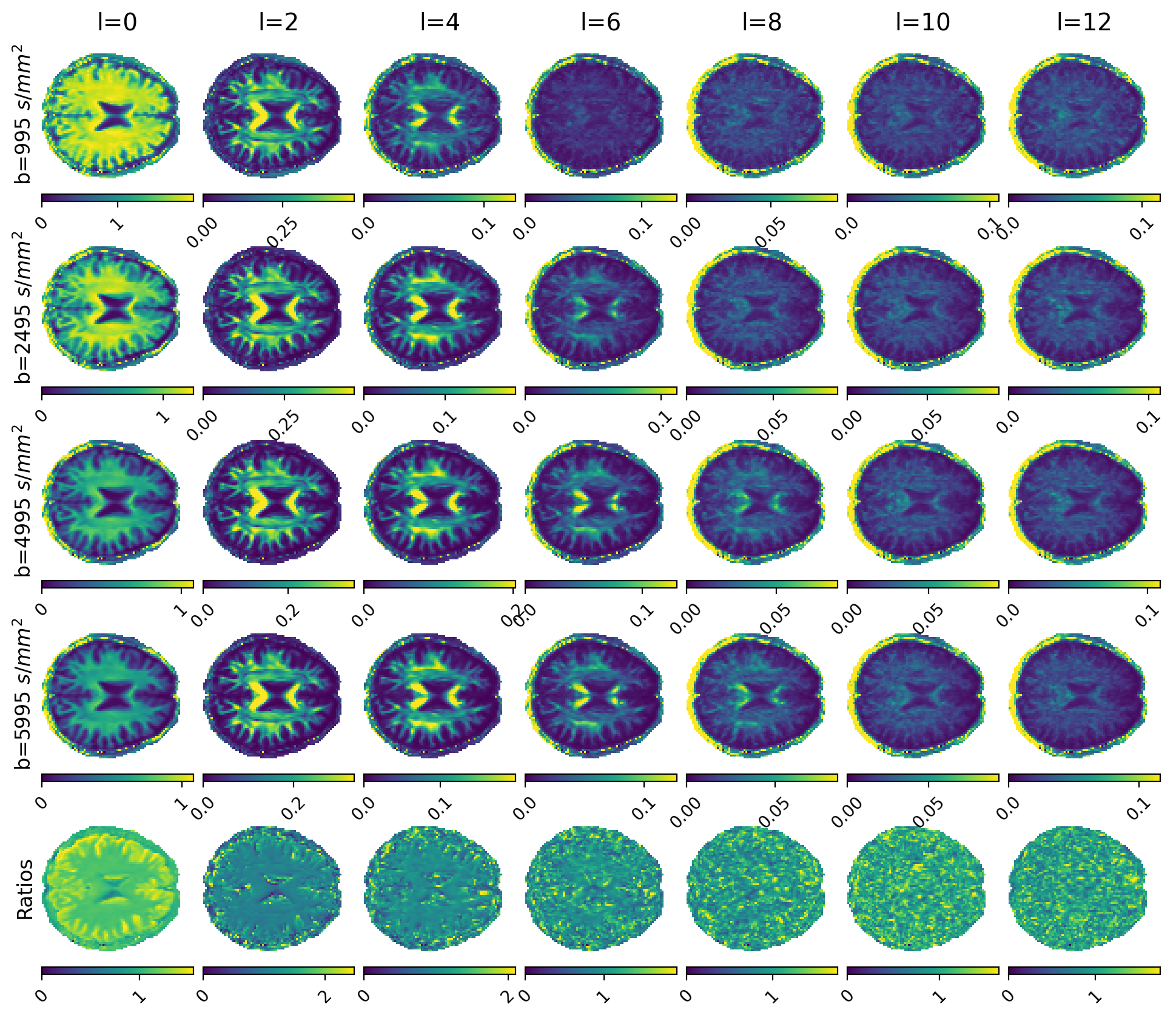
$$ \alpha_l(b_1, b_2, \lambda_{\parallel}, \lambda_{\bot}) := \sqrt{\frac{\Theta_l(b_1)}{\Theta_l(b_2)}} = |e^{(b_2-b_1)\lambda_{\bot}} \frac{\Psi_l(b_1 [\lambda_{\parallel} - \lambda_{\perp}])}{\Psi_l(b_2 [\lambda_{\parallel} - \lambda_{\perp}])}| $$
with $$$ \Psi_l$$$ defined in our work11 appendix A, and where
$$ \Theta_l(b) = \sum_{m=-l}^{l} c_{lm}(b)^2$$
are the l-band power spectra15. This calculation of $$$\alpha_l$$$ enables straightforward application of random forests (RF) regression (estimators=200, max-depth=20) between ratios (features) and diffusivities (labels). To account for noise in the data we calculated the ratios from noisy, simulated signals obtained with a multicompartmental model inclusive of WM, gray matter (GM), cellular (dot) fraction, and cerebrospinal fluid (CSF). WM includes intra- and extra-axonal compartments (axisymmetric tensors convolved with an orientation distribution function randomly selected from a custom database). GM and CSF are represented as isotropic diffusion. SH order was set to L=12. RF was trained using two million sets of seven ratios ($$$l=0,2,\dots,12$$$) and pairs of axial and radial diffusivity. Once diffusivities are estimated, we consider the powder average16
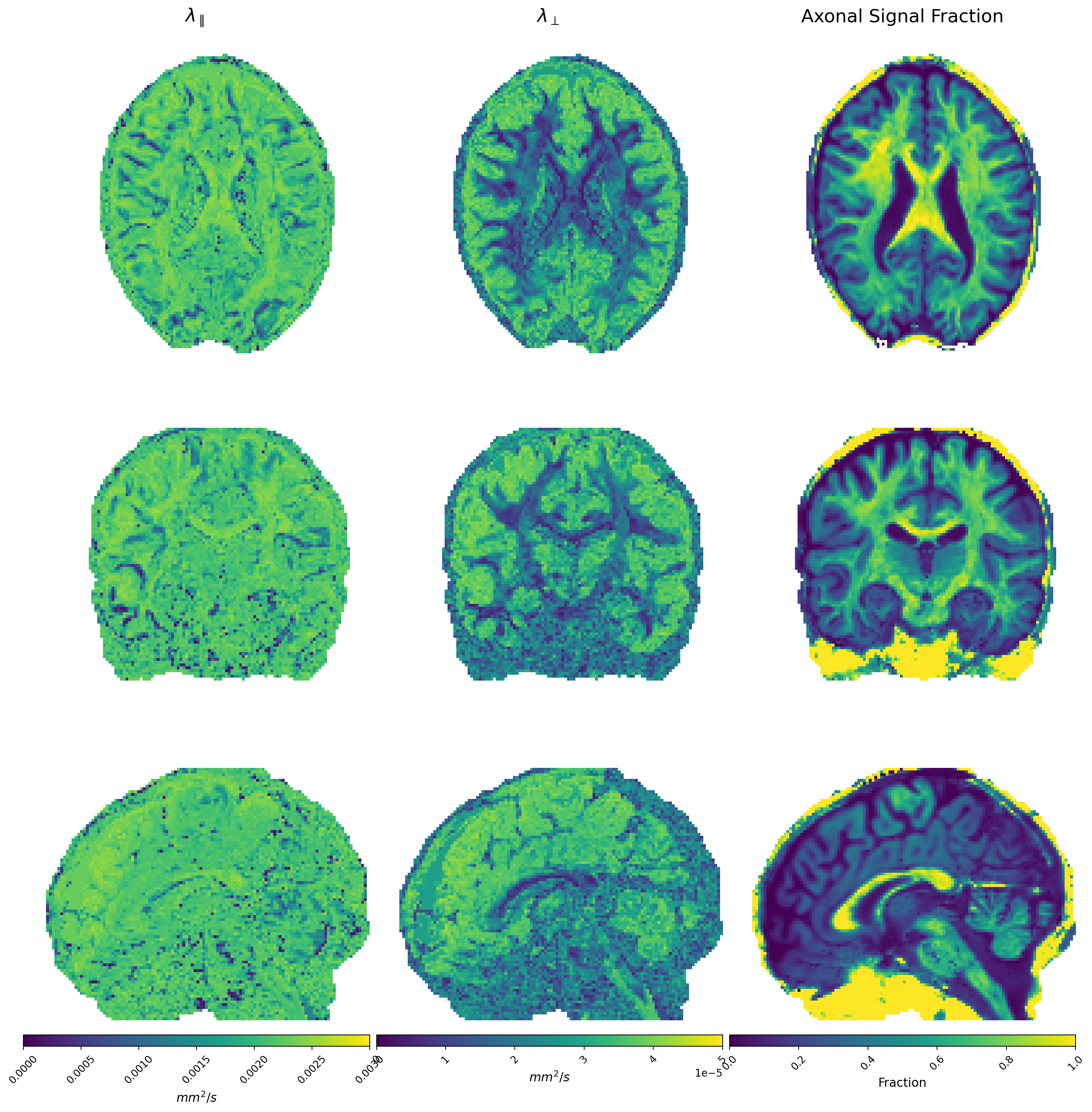
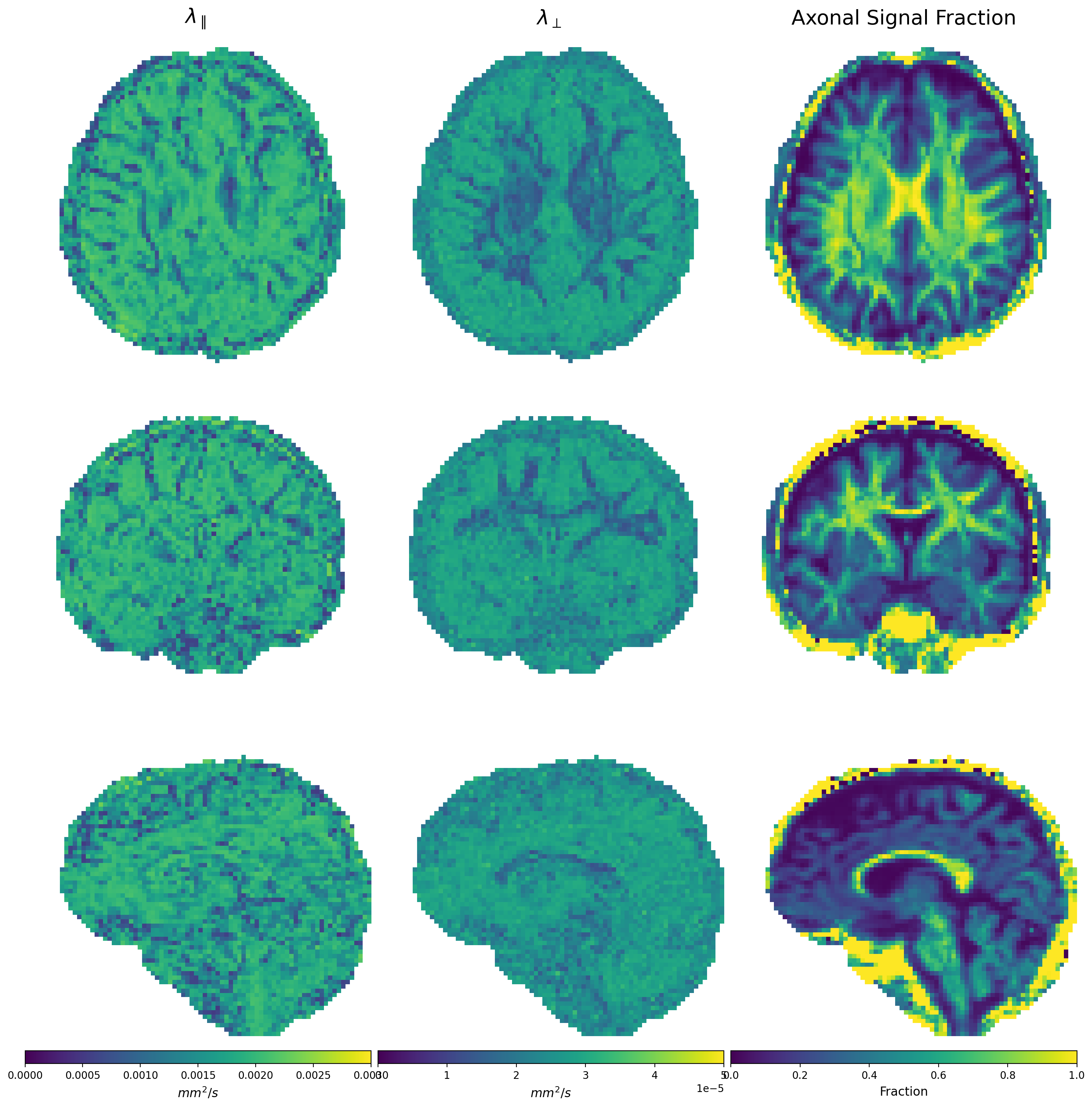
$$ \bar{S}(b) = Ce^{-b \lambda_{\bot}} \frac{\sqrt{\pi}}{2} \frac{\textrm{erf}\left[\sqrt{b(\lambda_{\parallel}-\lambda_{\bot})}\right]}{\sqrt{b(\lambda_{\parallel}-\lambda_{\bot})}}$$
from which we calculate the signal contribution from axons, $$$C$$$. The ASF is then obtained as $$$\textrm{ASF}=\frac{C}{S(b=0)}$$$.
Data
Clinical data was acquired from a healthy volunteer (34,male) on a Siemens Prisma 3T system collecting high b-values of ~5000 and ~6000 s/mm$$$^2$$$ (TE=98ms, TR = 3840ms, 2.3mm isotropic resolution, multi-band factor 3) while preclinical data comes from subject-1006 of the MGH Adult Diffusion HCP database with high b-values of ~5000 and ~10000 s/mm$$$^2$$$ (TE=57ms). Data was processed following a modified11 pipeline17 comprising denoising18 with Rician bias removal19, Gibbs ringing20, Eddy currents and motion21 correction.Results
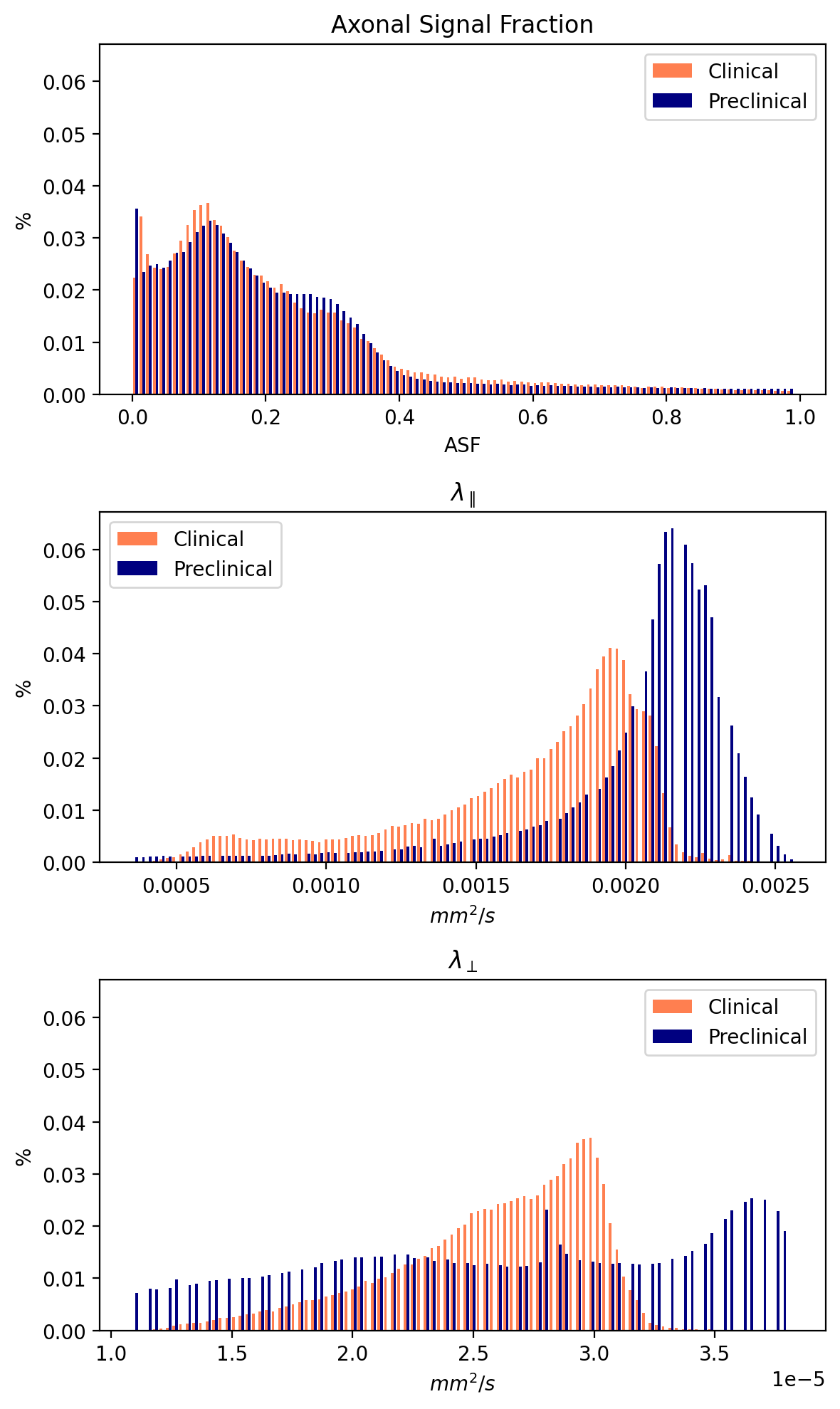
Figures 1 and 2 show $$$\Theta_l(b)$$$ for preclinical and clinical data with the high b-value ratios $$$\alpha_l(\cdot)$$$ at the bottom. Estimated diffusivities and ASF are shown in Figures 3 and 4 for preclinical and clinical data. Results show similar ASF patterns, with higher values in WM compared to GM, between clinical and preclinical data, indicating that the former yields qualitatively comparable values to the latter. However, we observe that clinical diffusivities occupy narrower ranges compared to the preclinical ones. Nevertheless, the ASF distributions we obtain are substantially equal (Fig. 5).Discussion
The adopted estimation strategy for diffusivities through l-band power spectra allowed us to obtain visually similar and comparable estimates between the clinical and the more sensitive (and high-SNR) preclinical data. Our method does not account yet for the known bias introduced by the presence of cells11. In feature work, we will try to exclude the bias to make unbiased estimates of diffusivities and ASF. To further assess the method, we should repeat multiple acquisitions on a Connectome scanner collecting preclinical and clinical-like data from the same subject.Conclusion
We have demonstrated the feasibility of estimating the ASF in both clinical and preclinical PGSE data.Acknowledgements
This work received funding from Danmarks Frie Forskningsfond (Denmark Free Research Foundation) with case number 3105-00129B. The acquisition of clinical data took place at Hvidovre Hospital (Denmark) and was authorized by the Danish ethical commission of Region Hovedstaden (Journal-nr.: H-21022514). Preclinical data were provided by the Human Connectome Project, WU-Minn Consortium (Principal Investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657) funded by the 16 NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by the McDonnell Center for Systems Neuroscience at Washington University.References
D.S. Novikov, E. Fieremans, S.N. Jespersen, V.G. Kiselev. Quantifying brain microstructure with diffusion MRI: theory and parameter estimation NMR Biomed. (2019), p. e3998.
Coronado-Leija, R., Abdollahzadeh, A., Lee, H. H., Coelho, S., Ades-Aron, B., Liao, Y., Salo, J.T, Tohka, J., Sierra, A., Novikov, D.S., & Fieremans, E. (2023). Volume electron microscopy in injured rat brain validates white matter microstructure metrics from diffusion MRI. arXiv preprint arXiv:2310.04608.
Jelescu I.O., Veraart J., Fieremans E., Novikov D.S. Degeneracy in model parameter estimation for multi-compartmental diffusion in neuronal tissue. NMR Biomed., 29 (1) (2016), pp. 33-47.
Kaden E., Kelm N.D., Carson R.P., Does M.D., Alexander D.C. Multi-compartment microscopic diffusion imaging NeuroImage, 139 (2016), pp. 346-359.
Fan Q., Nummenmaa A., Witzel T., Ohringer N., Tian Q., Setsompop K., Klawiter E.C., Rosen B.R., Wald L.L., Huang S.Y. Axon diameter index estimation independent of fiber orientation distribution using high-gradient diffusion MRI Neuroimage, 222 (2020), 117197.
Coelho, S., Pozo, J. M., Jespersen, S. N., Jones, D. K., & Frangi, A. F. (2019). Resolving degeneracy in diffusion MRI biophysical model parameter estimation using double diffusion encoding. Magnetic resonance in medicine, 82(1), 395-410.
Ramanna S., Moss H.G., McKinnon E.T., Yacoub E., Helpern J.A., Jensen J.H. Triple diffusion encoding MRI predicts intra-axonal and extra-axonal diffusion tensors in white matter. Magn. Reson. Med., 83 (6) (2020), pp. 2209-2220.
Reisert, M., Kiselev, V. G., & Dhital, B. (2019). A unique analytical solution of the white matter standard model using linear and planar encodings. Magnetic resonance in medicine, 81(6), 3819-3825.
Lundell, H., Nilsson, M., Dyrby, T., Parker, G., Cristinacce, P., Zhou, F.L., Topgaard, D., Lasič, S., 2019. Multidimensional diffusion MRI with spectrally modulated gradients reveals unprecedented microstructural detail. Scientific reports 9, 1–12.
Jensen J.H., Glenn G.R., Helpern J.A. Fiber ball imaging. Neuroimage, 124 (2016), pp. 824-833.
Pizzolato, M., Canales-Rodríguez, E. J., Andersson, M., & Dyrby, T. B. (2023). Axial and radial axonal diffusivities and radii from single encoding strongly diffusion-weighted MRI. Medical Image Analysis, 86, 102767.
Veraart J., Fieremans E., Novikov D.S. On the scaling behavior of water diffusion in human brain white matter. NeuroImage, 185 (2019), pp. 379-387.
Descoteaux M., Angelino E., Fitzgibbons S., Deriche R. Regularized, fast, and robust analytical Q-ball imaging. Magn. Reson. Med., 58 (3) (2007), pp. 497-510.
Anderson A.W. Measurement of fiber orientation distributions using high angular resolution diffusion imaging. Magn. Reson. Med. Off. J. Int. Soc. Magn. Reson. Med., 54 (5) (2005), pp. 1194-1206.
Reisert M., Kellner E., Dhital B., Hennig J., Kiselev V.G. Disentangling micro from mesostructure by diffusion MRI: A Bayesian approach. Neuroimage, 147 (2017), pp. 964-975.
Kroenke C.D., Ackerman J.J., Yablonskiy D.A.On the nature of the NAA diffusion attenuated MR signal in the central nervous system. Magn. Reson. Med. Off. J. Int. Soc. Magn. Reson. Med., 52 (5) (2004), pp. 1052-1059.
Ma X., Uğurbil K., Wu X. Denoise magnitude diffusion magnetic resonance images via variance-stabilizing transformation and optimal singular-value manipulation. Neuroimage, 215 (2020), Article 116852.
Gavish M., Donoho D.L. Optimal shrinkage of singular values. IEEE Trans. Inform. Theory, 63 (4) (2017), pp. 2137-2152.
Foi A. Noise estimation and removal in MR imaging: The variance-stabilization approach2011. IEEE International Symposium on Biomedical Imaging: From Nano to Macro, IEEE (2011), pp. 1809-1814.
Kellner E., Dhital B., Kiselev V.G., Reisert M. Gibbs-ringing artifact removal based on local subvoxel-shifts. Magn. Reson. Med., 76 (5) (2016), pp. 1574-1581.
Andersson J.L.R., Sotiropoulos S.N. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. Neuroimage, 125 (2016), pp. 1063-1078.
Figures