3465
Time-Dependent Standard Model of diffusion in human brain white matter evaluated in vivo on the high gradient performance Connectome 2.0 scanner1Athinoula A. Martinos Center for Biomedical Imaging, Charlestown, MA, United States, 2Department of Radiology, Harvard Medical School, Boston, MA, United States, 3Center for Biomedical Imaging, Department of Radiology, New York University School of Medicine, New York, NY, United States, 4Center for Advanced Imaging Innovation and Research (CAI2R), Department of Radiology, New York University School of Medicine, New York, NY, United States
Synopsis
Keywords: Microstructure, Microstructure
Motivation: While most biophysical models in brain white matter estimate Gaussian compartment parameters, characteristic length scales of tissue microstructure can only be obtained from non-Gaussian features.
Goal(s): To introduce time dependence into the Standard Model of diffusion; to in vivo evaluate non-Gaussian signatures of diffusion in intra- and extra-neurite spaces, irrespective of neurite orientation dispersion.
Approach: We perform diffusion measurements on the Connectome 2.0 scanner in healthy volunteers at short times (13-30 ms) and estimate time-dependent diffusion parameters using GPU-accelerated fitting.
Results: Time-dependent diffusion signals up to 2nd-order in spherical harmonics provide sensitivity potentially related to axonal beadings and packing correlation length.
Impact: We demonstrated the feasibility of mapping time-dependent diffusion in human white matter in vivo using the Connectome 2.0 scanner. This potentially provides novel biomarkers sensitive to axon beadings and packing length scales for investigation of neurological disorders.
Introduction
In the standard model of diffusion in white matter (WM)1-2, signal is modelled as the convolution of an orientation distribution function (ODF) and kernel/response of a fiber fascicle represented by Gaussian compartments. However, the axonal microstructure suggests non-Gaussian (e.g., time-dependence) diffusion in each compartment, as observed previously in brain WM3-10.Here, we (i) introduce the asymptotic relation of diffusivity and kurtosis time-dependence in each compartment in tortuosity limit into the standard model(Fig.1)9, (ii) fit this Time-Dependent Standard Model (TDSM) to in vivo diffusion measurements in 10 subjects scanned on Connectome 2.0 at relatively shorter times (13-30ms) and moderate b-value$$$\leq$$$2,400s/mm2(Fig.1b), and (iii) estimate time-dependent diffusion parameters using an GPU-accelerated fitting pipeline11-12, offering potential biomarkers for beading and packing length scales.
Theory
We introduce diffusion Time-dependence into the standard model in neuronal tissues9(Fig.1a). The diffusivity and kurtosis Time-dependence (up to ~b2) in intra- and extra-neurite spaces provide the asymptotic form of diffusion signals at moderate diffusion weighting b in tortuosity (long time) regime3-10:$$D_a(t)=D_{a,\infty}+c_a\cdot\frac{1}{\sqrt{t}}\,,$$
$$K_a(t)=\frac{2c_a}{D_{a,\infty}}\cdot\frac{1}{\sqrt{t}}\,,$$
$$D_e^\parallel(t)=D_{e,\infty}^\parallel+c_e^\parallel\cdot\frac{1}{\sqrt{t}}\,,$$
$$K_e^\parallel(t)=\frac{2c_e^\parallel}{D_{e,\infty}^\parallel}\cdot\frac{1}{\sqrt{t}}\,,$$
$$D_e^\perp(t)=D_{e,\infty}^\perp+c_e^\perp\cdot\frac{\log{\Delta/\delta}+3/2}{\Delta-\delta/3}\,,$$
$$K_e^\perp=\frac{6c_e^\perp}{D_{e,\infty}^\perp}\cdot\frac{\log{\Delta/\delta}+3/2}{\Delta-\delta/3}\,.$$
Time-dependence is parameterized by the inter-pulse duration $$$\Delta$$$ and pulse width $$$\delta$$$. The ideal diffusion time $$$t$$$ is a function of $$$\Delta$$$ and $$$\delta$$$ due to the filtering effect4,10 of pulsed-gradients. To increase the precision of parameter fitting and avoid parameter degeneracy in the standard model1,13, we assume that the diffusivity and kurtosis parallel to axons are the same in intra- and extra-neurite spaces ($$$D_{e,\infty}^\parallel=D_{a,\infty}$$$, $$$c_e^\parallel=c_a$$$). The time-dependencies transverse to axons are characterized by the diffusivity $$$D_e^\perp(t)$$$ and kurtosis $$$K_e^\perp(t)$$$ transverse to axons in the extra-neurite space.
Tissue parameters include the intra-neurite volume fraction $$$f$$$, bulk diffusivity $$$D_{a,\infty},D_{e,\infty}^\perp$$$ in $$$t\to\infty$$$ limit, strengths of restrictions $$$c_a,c_e^\perp$$$ in each compartment and direction. We denote $$$x=\{f,D_{a,\infty},D_{e,\infty}^{\perp}, c_{a}, c_{e}^{\perp}\}$$$ as the parameter set (Fig.1a).
We numerically decompose time-dependent signal kernels using the spherical harmonics for each b-value and calculate rotational invariants ($$$S_{l=0}(b),S_{l=2}(b)$$$)1,14 up to order $$$l=2$$$. We estimate all parameters without applying any assumptions of ODF, whose 2nd order rotational invariant $$$p_2\in$$$[0,1] is estimated.
Methods
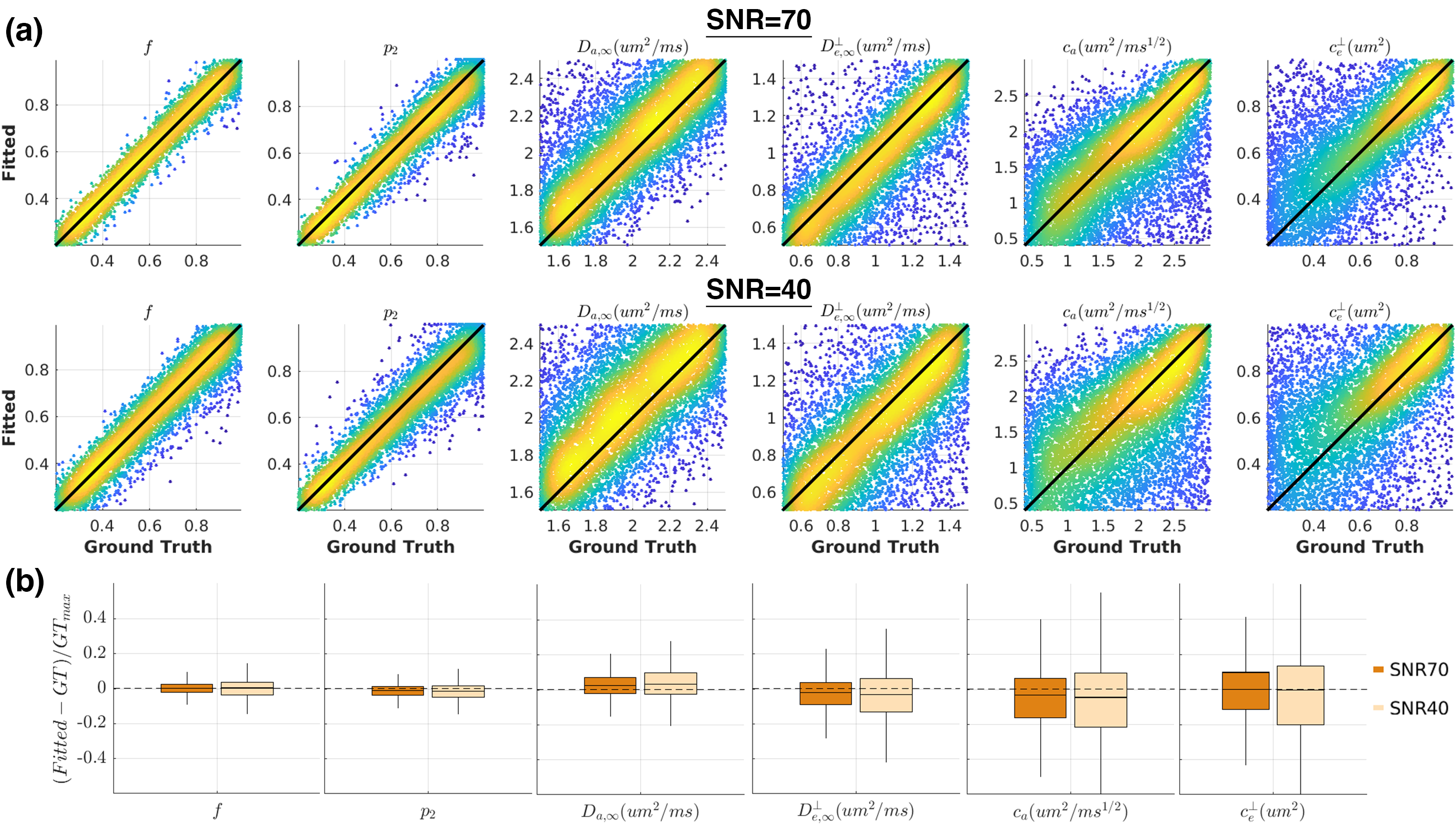
Noise propagationWe evaluated the performance of TDSM fitting using the GPU-accelerated ‘askAdam’ framework11-12. We generated diffusion signals with the acquisition protocol in Fig.1b. We randomly chose tissue parameters in 10,000 different combinations within the following range: $$$f$$$=[0.2,0.99], $$$D_{a,\infty}$$$=[1.5,2.5]μm2/ms, $$$D_{e,\infty}^\perp$$$=[0.5,1.5]μm2/ms, $$$c_a$$$=[0.4,3]μm2/ms1/2, $$$c_e^\perp$$$=[0.2,1]μm2, $$$p_2$$$=[0.2,1]. We added Gaussian noise to simulate signals at SNR=70 and SNR=40 and fitted TDSM to the simulated signals.
In vivo MRI
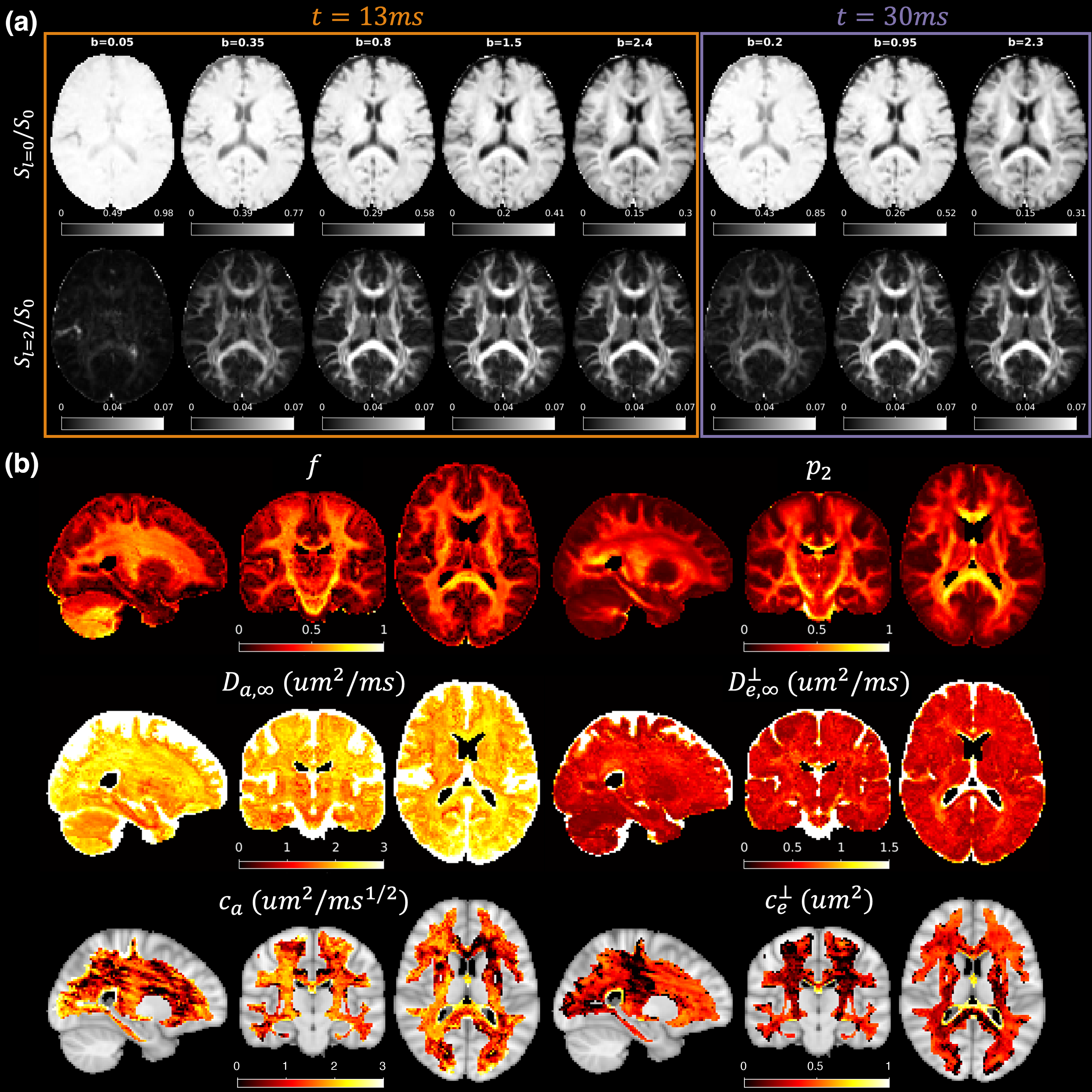
We performed diffusion MRI in 10 healthy subjects (9F1M, 26-49years) on the 3T Connectome 2.0 scanner (MAGNETOM Connectom.X, Siemens Healthineers) using a 72-channel head coil15. We acquired an MPRAGE of 0.9mm isotropic resolution and dMRI with the following protocol: GRAPPA=2, SMS=2, 2mm isotropic, PF=6/8, TR/TE=3600/54ms using the diffusion scheme in Fig.1b, with interspersed b=0 images acquired every 16 DWIs. The total scan time of low b-value data used in this study (b$$$\leq$$$2,400s/mm2) is 20 min. DWIs were processed using DESIGNER pipeline16.
ROI analysis
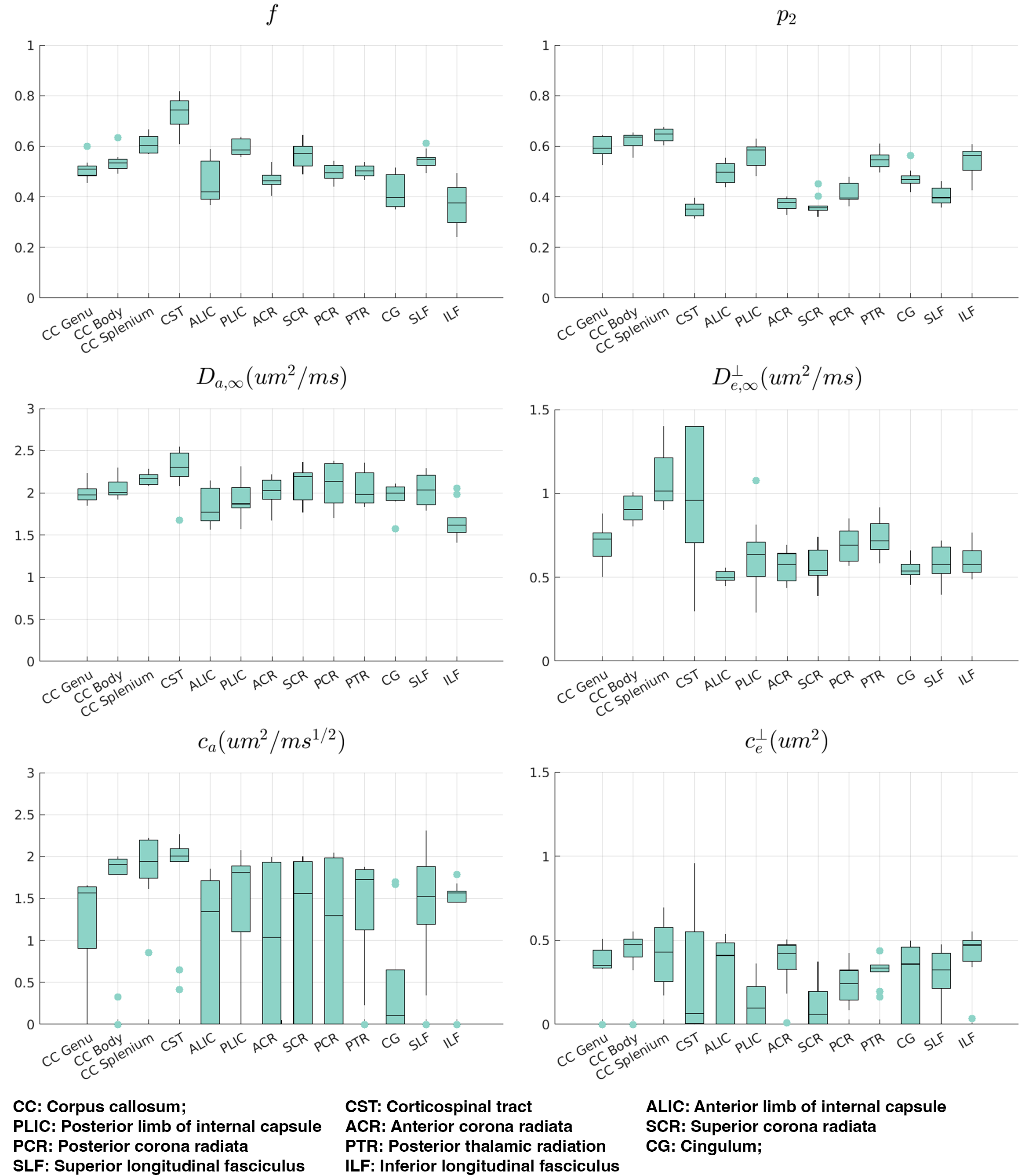
We computed signal-rotational invariants ($$$l=0,2$$$) in each voxel and averaged them over WM ROIs in JHU-DTI WM atlas17. We fitted TDSM to averaged signal rotational invariants in each ROI with constraints $$$\{f,p_2\}\in$$$[0,1], $$$\{D_{a,\infty},D_{e,\infty}^\perp\}\in$$$[0,3]µm2/ms, $$$c_a\in$$$[0,4]µm2/ms1/2, and $$$c_e^\perp\in$$$[0,2]µm2.
TDSM parameter mapping
Signal rotational invariants were transformed to the MNI152-2mm space and averaged across all subjects. Parameters were estimated by fitting TDSM to signal rotational invariants using ‘askAdam’.
Results
All TDSM parameters show low estimation bias (<5%) that does not increase with lower SNR, though the precision is reduced(Fig.2). $$$c_{a}$$$ and $$$c_{e}^{\perp}$$$ are the most challenging parameters to estimate especially when their values are low(Fig.2a) with interquartile range (IQR) $$$c_{a}$$$=31% and IQR $$$c_{e}^{\perp}$$$=34% at SNR=40(Fig.2b).We observed higher inter-subject variabilities on $$$c_{a}$$$ and $$$c_{e}^{\perp}$$$ compared to other TDSM metrics across all WM ROIs on the vivo data. All WM ROIs exhibit certain degrees of longitudinal time-dependence (median>1µm2/ms1/2) except for the cingulum(Fig.3), and the corticospinal tracts only show weak transverse time-dependence.
On the $$$c_{e}^{\perp}$$$ map estimated using group-averaged signal rotational invariants, low values are clearly shown along the superior corona radiata (SCR), posterior limb of the internal capsule (PLIC) and splenium of CC. SCR and PLIC also show relatively stronger longitudinal time-dependencies on the $$$c_{a}$$$ map compared to their surroundings WM tissue.
Discussion and Conclusions
The high inter-subject variability of $$$c_{a}$$$ and $$$c_{e}^{\perp}$$$ can be caused by the noisy estimation shown in noise propagations. Our $$$c_{a}$$$ values are generally lower than those previously reported at longer times (t=45-600ms)5. Future work will focus on optimising the acquisition protocol to improve the stability of the time-dependent parameter estimation for TDSM mapping at the individual level and investigating the association between structural disorders and the time-dependent diffusion parameters.Acknowledgements
This study is support by NIH under the award number: DP5OD031854, R01NS118187, P41EB015896, P41EB030006, U01EB026996, S10RR023401, S10RR019307, R21NS081230, R01NS088040, P41EB017183.References
1. Novikov DS, Veraart J, Jelescu IO, Fieremans E. Rotationally-invariant mapping of scalar and orientational metrics of neuronal microstructure with diffusion MRI. NeuroImage. 2018;174:518-538.
2. Novikov DS, Fieremans E, Jespersen SN, Kiselev VG. Quantifying brain microstructure with diffusion MRI: Theory and parameter estimation. NMR in Biomedicine. 2019 Apr;32(4):e3998.
3. Novikov DS, Jensen JH, Helpern JA, Fieremans E. Revealing mesoscopic structural universality with diffusion. Proc Natl Acad Sci. 2014;111(14):5088-5093.
4. Burcaw LM, Fieremans E, Novikov DS. Mesoscopic structure of neuronal tracts from time-dependent diffusion. NeuroImage. 2015;114:18-37.
5. Fieremans E, Burcaw LM, Lee HH, Lemberskiy G, Veraart J, Novikov DS. In vivo observation and biophysical interpretation of time-dependent diffusion in human white matter. NeuroImage. 2016;129:414-427.
6. Jespersen SN, Olesen JL, Hansen B, Shemesh N. Diffusion time dependence of microstructural parameters in fixed spinal cord. Neuroimage. 2018 Nov 15;182:329-42.
7. Lee HH, Fieremans E, Novikov DS. What dominates the time dependence of diffusion transverse to axons: Intra-or extra-axonal water?. NeuroImage. 2018 Nov 15;182:500-10.
8. Lee HH, Papaioannou A, Kim SL, Novikov DS, Fieremans E. A time-dependent diffusion MRI signature of axon caliber variations and beading. Communications biology. 2020 Jul 7;3(1):354.
9. Lee HH, Fieremans E, Novikov DS. LEMONADE (t): exact relation of time-dependent diffusion signal moments to neuronal microstructure. In: Proceedings of the 26th Annual Meeting of ISMRM, Paris, France 2018.
10. Lee, HH, Tian Q., Sheft M., et al. (2023). The influence of axonal beading and undulation on axonal diameter mapping. bioRxiv.
11. Chan KS, Kim TH, Bilgic B, Marques JP. Semi-supervised learning for fast multi-compartment relaxometry myelin water imaging (MCR-MWI). In: Proceedings 30th. Annual Meeting International Society for Magnetic Resonance in Medicine. London, United Kingdom.
12. Marques JP, Hollander DD, Chan KS. Improved R2* and QSM mapping for dummies - ask Adam. In: Proceedings 31. Annual Meeting International Society for Magnetic Resonance in Medicine. Toronto, Canada.
13. Jelescu IO, Veraart J, Fieremans E, Novikov DS. Degeneracy in model parameter estimation for multi-compartmental diffusion in neuronal tissue. Nmr Biomed. 2015;29(1):33-47.
14. Reisert, M., Kellner, E., Dhital, B., Hennig, J., & Kiselev, V. G. (2017). Disentangling micro from mesostructure by diffusion MRI: a Bayesian approach. NeuroImage, 147, 964-975.
15. Mahmutovic M, Shrestha M, Ramos-Llordén G, et al. A 72-channel Head Coil with an Integrated 16-Channel Field Camera for the Connectome 2.0 Scanner. Submitted to ISMRM 2024.
16. Ades-Aron B, Veraart J, Kochunov P, et al. Evaluation of the accuracy and precision of the diffusion parameter EStImation with Gibbs and NoisE removal pipeline. NeuroImage. 2018;183:532-543.
17. Mori S, Wakana S, Van Zijl PC, Nagae-Poetscher LM. MRI atlas of human white matter. Elsevier; 2005 May 11.
Figures