3463
Water exchange as measured by diffusion MRI with free gradient waveforms: A potential biomarker of dendritic spine morphology1Medical Radiation Physics, Lund, Lund University, Lund, Sweden, 2Cardiff University Brain Research Imaging Centre (CUBRIC), School of Psychology, Cardiff University, Cardiff, United Kingdom, 3School of Computer Science and Informatics, Cardiff University, Cardiff, United Kingdom, 4F.M. Kirby Research Center for Functional Brain Imaging, Kennedy Krieger Institute, Baltimore, MD, United States, 5Department of Neurology, Johns Hopkins University School of Medicine, Baltimore, MD, United States, 6Department of Clinical Sciences Lund, Radiology, Lund University, Lund, Sweden
Synopsis
Keywords: Microstructure, Microstructure, Exchange
Motivation: Time-dependent diffusion MRI reveals fast exchange in grey matter, but it remains unclear whether permeative or non-permeative exchange explains these findings.
Goal(s): We set to investigate the effect of non-permeative exchange induced by dendritic spines on the time-dependence of the diffusion-weighted signal and the exchange estimates obtained from the signal.
Approach: Monte Carlo simulations were performed in synthetic dendrites with varying spine densities and signals were generated using free gradient waveforms. The intracellular signals were analysed using a restriction-exchange framework from previous work.
Results: Dendritic spines give the same signal time-dependence as permeative exchange. Estimated exchange rates increase with spine density.
Impact: Dendritic spines may explain the exchange rates observed with diffusion MRI in grey matter. Furthermore, exchange measurements provide a potential biomarker of dendritic spine morphology, which is important because spine density is implicated in, for example, learning and psychiatric disorders.
Introduction
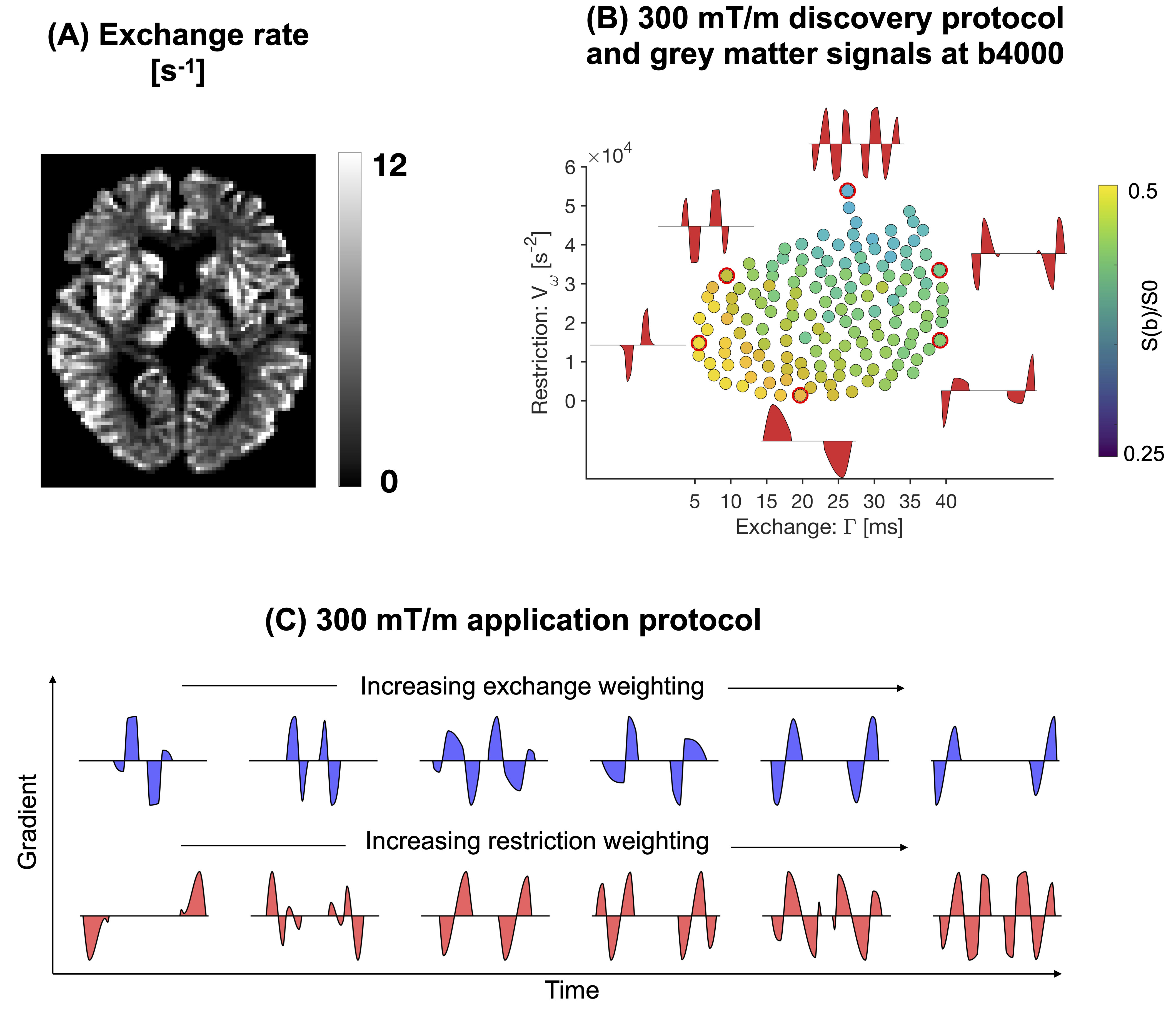
Time-dependent diffusion MRI enables estimation of water exchange in biological tissue1,2. Recent work has shown that complex geometries can induce apparent water exchange without membrane permeation3. Here, we investigate this mode of exchange using Monte Carlo simulations in synthetic dendrites featuring varying spine densities4. We use the framework presented in ref5 to define gradient waveforms efficient for detection of water exchange, as well as for the analysis of the simulated intracellular signals. Results show that the estimated exchange rates are a function of the spine density.Theory
The influence of restricted diffusion and exchange on the diffusion-weighted signal is described by5$$\ln(S/S_0)\approx-b\cdot[E_{\beta_0}+V_{\omega}E_{\beta_2}]+\frac{1}{2}b^2\cdot[V_{\beta_0}+2V_{\omega}C_{\beta_0\beta_2}+V_{\omega}^2V_{\beta_2}]\cdot(1-k\Gamma),\hspace{24pt}(1)$$
where$$$\hspace{2pt}S_0\hspace{2pt}$$$denotes the non-diffusion-weighted signal, three parameters describe the experiment ($$$b,V_{\omega}$$$, and $$$\Gamma$$$), and six parameters describe the microstructure ($$$E_{\beta_0}, E_{\beta_2},V_{\beta_0},C_{\beta_0\beta_2},V_{\beta_2}\hspace{2pt}$$$and$$$\hspace{2pt}k$$$). Of the experiment parameters,$$$\hspace{2pt}b\hspace{2pt}$$$is the b-value given by$$$\hspace{2pt}b=\int_0^Tq(t)^2dt\hspace{2pt}$$$where$$$\hspace{2pt}q(t)=\gamma \int_0^t g(\tau)d\tau\hspace{2pt}$$$is the dephasing q-vector,$$$\hspace{2pt}g(\tau)\hspace{2pt}$$$is the gradient waveform and$$$\hspace{2pt}\gamma\hspace{2pt}$$$is the gyromagnetic ratio,$$$\hspace{2pt}V_{\omega}\hspace{2pt}$$$is the restriction-weighting given by
$$V_{\omega}=\frac{1}{2\pi b}\int_{-\infty}^{\infty}|q(\omega)|^2d\omega=\frac{\gamma^2}{b}\int_0^Tg^2(t)dt,\hspace{24pt}(2)$$
where$$$\hspace{2pt}q(\omega)\hspace{2pt}$$$is the Fourier transform of$$$\hspace{2pt}q(t)\hspace{2pt}$$$.$$$\hspace{2pt}\Gamma\hspace{2pt}$$$is the exchange-weighting defined as
$$\Gamma=2\int_0^Tt\tilde{q_4}(t)dt,\hspace{24pt}(3)$$
where$$$\hspace{2pt}\tilde{q_4}{(t)}=1/b^2\cdot\int_0^Tq^2(t')q^2(t'+t)dt'$$$. Note that,$$$\hspace{2pt}E_{\beta_0}=\langle\beta_0\rangle\hspace{2pt}$$$is the average time-independent diffusivity,$$$\hspace{2pt}E_{\beta_2}=\langle\beta_2\rangle\hspace{2pt}$$$is the average restriction coefficient,$$$\hspace{2pt}V_{\beta_0}\hspace{2pt}$$$and$$$\hspace{2pt}V_{\beta_2}\hspace{2pt}$$$are the variances in$$$\hspace{2pt}\beta_0\hspace{2pt}$$$and$$$\hspace{2pt}\beta_2\hspace{2pt}$$$, while$$$\hspace{2pt}C_{\beta_0\beta_2}\hspace{2pt}$$$is the covariance between them and, finally,$$$\hspace{2pt}k\hspace{2pt}$$$is the exchange rate. By defining a set of gradient waveforms that independently probe the parameters$$$\hspace{2pt}\Gamma\hspace{2pt}$$$and$$$\hspace{2pt}V_{\omega}\hspace{2pt}$$$, restriction- and exchange-related parameters can be reliably estimated simultaneously.
Methods
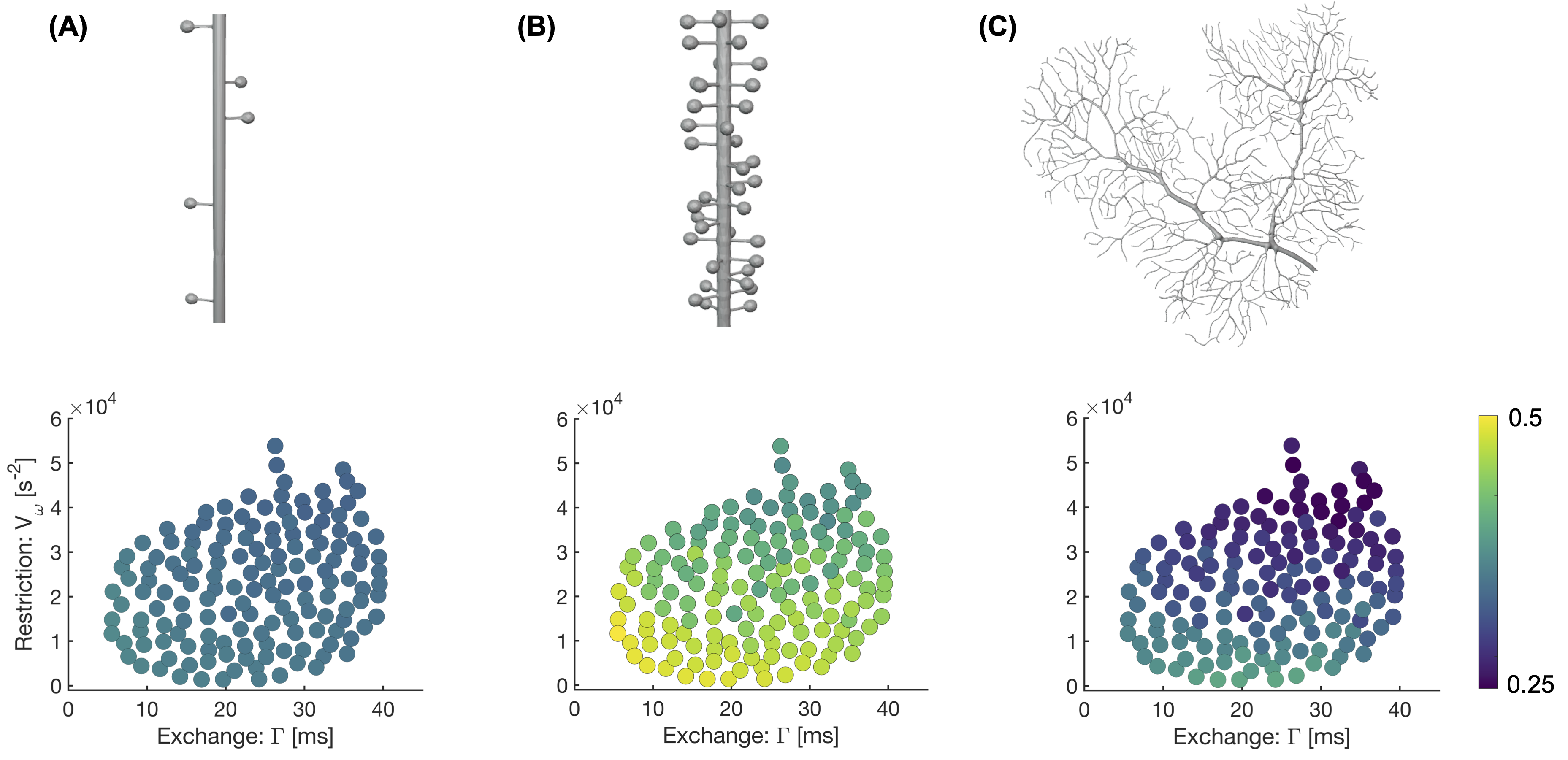
Numerical simulationsMonte Carlo simulations were performed to investigate how the intracellular signal and the estimated exchange rates depend on spine density. Substrates mimicking dendrites were constructed following Palombo et al.4: cylinders lined with identical randomly positioned spines of different densities between 0 and 1 µm-1 (cf. Fig.2). The cylinders had a diameter of 1 µm and the spines had head diameters of 1 µm and neck diameters of 0.25 µm. For comparison, simulations were also run in the dendritic tree from a Purkinje cell segmented from the mouse brain (average dendrite diameter of 1.27 µm and no spines). In all simulations, only intracellular spins were considered. Simulation parameters were: number of particles = 200k, simulation time-step = 0.2 µs, bulk diffusivity = 2 µm2/ms and total simulation time = 110 ms. Signals were generated using the free gradient waveforms (FWF) shown in Fig.1 and a single-diffusion-encoding protocol with varying diffusion time. Signals from all waveforms were averaged across 20 encoding directions.
Data analysis
The signals were analysed by visualisation in the restriction-exchange landscape shown in Fig.1B to determine the dominant mechanism of time-dependence. Exchange estimates were obtained by fitting Eq.1. Following previous work6, the fitting replaced the factor$$$\hspace{2pt}h(k)=1-k\Gamma\hspace{2pt}$$$in Eq.1 with the more accurate$$$\hspace{2pt}h(k)=2\int_0^T\exp(-kt)\tilde{q_4}(t)dt$$$.
Results
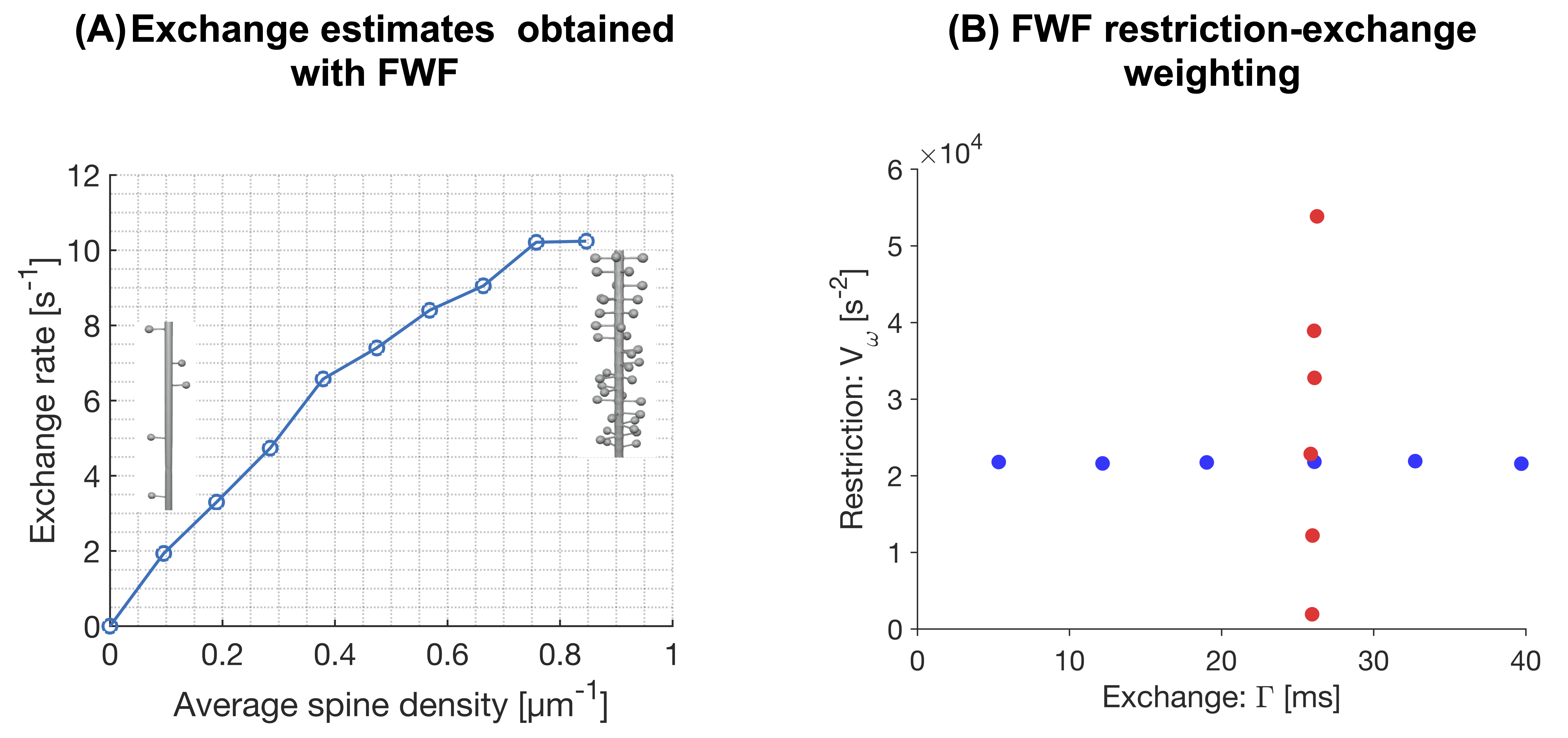
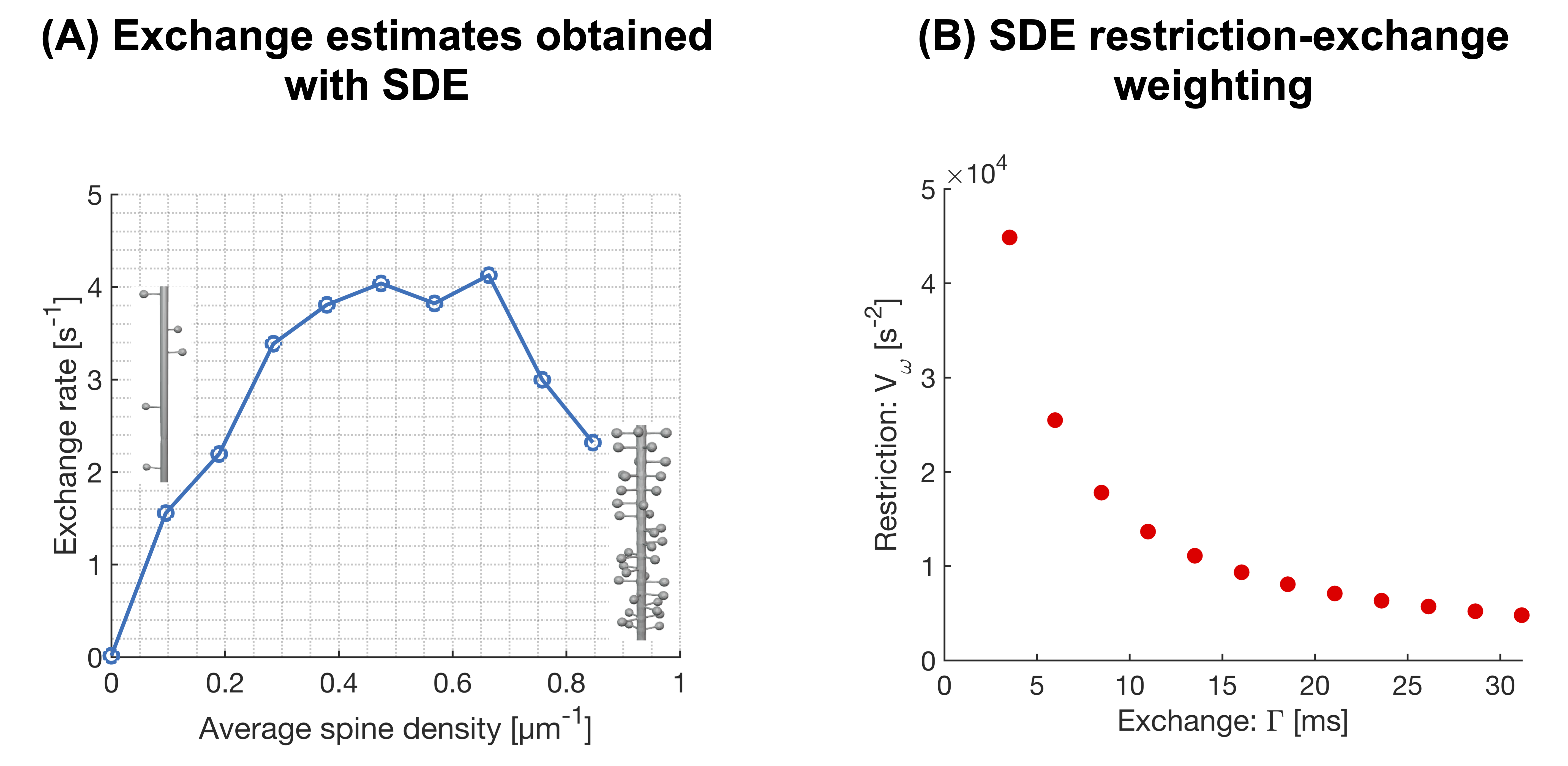
Figure 2 shows powder-averaged intracellular signals at b = 4000 s/mm2 obtained with the protocol in Fig.1B in three different substrates. At low spine densities (panel A), there is no clear trend. At higher spine densities (B), both restriction and exchange are important mechanisms of time dependence (signal variation along both x- and y-axes). Time dependence in the dendritic tree of the Purkinje cell is dominated by restriction.Figure 3A shows exchange estimates obtained by fitting Eq.1 to intracellular signals generated with the protocol in Fig.1C. The estimated exchange rates increase with spine density. Corresponding estimates obtained with a single-diffusion-encoding protocol with varying diffusion times are shown in Fig.4A, where a non-monotonic dependence of the exchange rate on spine density is evident. The discrepancy arises because free waveforms enable an independent variation of the restriction-and exchange encoding (Fig.3B), while single-diffusion-encoding gives a strong correlation between the two encoding axes (Fig.4B).
Discussion
This work demonstrates that the intracellular diffusion-weighted signal in spiny structures exhibits time-dependence signatures consistent with both restricted diffusion and exchange. The estimated exchange times decreased with spine density down to approximately 100 ms, which is within the literature range7,8. We note that this non-permeative mode of exchange is indistinguishable from permeative exchange (they both manifest in the signal as a temporal loss of diffusional heterogeneity), which creates a challenge in relating the exchange rate estimated from diffusion-weighted signals to cell permeability. Furthermore, this mechanism of exchange may explain the relatively fast exchange rates reported in cortical grey matter6,8 (cf. Fig1A). The mechanism may also account for the fast exchange rates reported in the cerebellar cortex6, where Purkinje cells give a high concentration of spiny dendrites in the molecular layer9.Limitations of this work are associated with the analysis framework, which accounts for exchange and restricted diffusion but ignores structural disorder10, anisotropy11,12 and microscopic kurtosis13. Another limitation is that we did not consider diffusion in the extracellular space.
There are numerous potential applications of this approach, as spine density is implicated in learning, development, degeneration, as well as in various psychiatric disorders.
Acknowledgements
No acknowledgement found.References
1. Kärger J. NMR self-diffusion studies in heterogeneous systems. Advances in Colloid and Interface Science. 1985;23:129-148. doi:10.1016/0001-8686(85)80018-X
2. Reynaud O. Time-Dependent Diffusion MRI in Cancer: Tissue Modeling and Applications. Front Phys. 2017;5. doi:10.3389/fphy.2017.00058
3. Khateri M, Reisert M, Sierra A, Tohka J, Kiselev VG. What does FEXI measure? NMR in Biomedicine. 2022;35(12):e4804. doi:10.1002/nbm.4804
4. Palombo M, Ligneul C, Hernandez-Garzon E, Valette J. Can we detect the effect of spines and leaflets on the diffusion of brain intracellular metabolites? NeuroImage. 2018;182:283-293. doi:10.1016/j.neuroimage.2017.05.003
5. Chakwizira A, Westin CF, Brabec J, et al. Diffusion MRI with pulsed and free gradient waveforms: Effects of restricted diffusion and exchange. NMR in Biomedicine. 2022;n/a(n/a):e4827. doi:10.1002/nbm.4827
6. Chakwizira A, Zhu A, Foo T, Westin CF, Szczepankiewicz F, Nilsson M. Diffusion MRI with free gradient waveforms on a high-performance gradient system: Probing restriction and exchange in the human brain. NeuroImage. 2023;283:120409. doi:10.1016/j.neuroimage.2023.120409
7. Jelescu IO, Palombo M, Bagnato F, Schilling KG. Challenges for biophysical modeling of microstructure. Journal of Neuroscience Methods. 2020;344:108861. doi:10.1016/j.jneumeth.2020.108861
8. Olesen JL, Østergaard L, Shemesh N, Jespersen SN. Diffusion time dependence, power-law scaling, and exchange in gray matter. NeuroImage. 2022;251:118976. doi:10.1016/j.neuroimage.2022.118976
9. Voogd J, Glickstein M. The anatomy of the cerebellum. Trends Neurosci. 1998;21(9):370-375. doi:10.1016/s0166-2236(98)01318-6
10. Novikov DS, Jensen JH, Helpern JA, Fieremans E. Revealing mesoscopic structural universality with diffusion. Proceedings of the National Academy of Sciences. 2014;111(14):5088-5093. doi:10.1073/pnas.1316944111
11. Lasič S, Yuldasheva N, Szczepankiewicz F, et al. Stay on the Beat With Tensor-Valued Encoding: Time-Dependent Diffusion and Cell Size Estimation in ex vivo Heart. Frontiers in Physics. 2022;10. Accessed May 19, 2022. https://www.frontiersin.org/article/10.3389/fphy.2022.812115
12. Lundell H, Nilsson M, Dyrby TB, et al. Multidimensional diffusion MRI with spectrally modulated gradients reveals unprecedented microstructural detail. Sci Rep. 2019;9(1):9026. doi:10.1038/s41598-019-45235-7
13. Henriques RN, Jespersen SN, Shemesh N. Evidence for microscopic kurtosis in neural tissue revealed by correlation tensor MRI. Magnetic Resonance in Medicine. 2021;86(6):3111-3130. doi:10.1002/mrm.28938
Figures