3462
Incorporating mesoscopic orientation dependent R2 from magnetic susceptibility into the Standard Model of Diffusion in White Matter1Center of Functionally Integrative Neuroscience, Department of Clinical Medicine, Aarhus University, Aarhus, Denmark, 2Champalimaud Research,Champalimaud Centre for the Unknown, Lisbon, Portugal, 3Division of Medical Physics, Department of Radiology, University Medical Center Freiburg, Freiburg, Germany, 4Department of Physics and Astronomy, Aarhus University, Aarhus, Denmark
Synopsis
Keywords: Microstructure, Microstructure
Motivation: $$$R_2$$$ in WM is orientation dependent due to microscopic magnetic anisotropy. So far, the Standard Model of diffusion (SM) has been extended to only include isotropic $$$R_2$$$ (TEdDI).
Goal(s): Our goal is to characterize $$$R_2$$$ anisotropy of a multi-echo dMRI signal for long diffusion times and incorporate $$$R_2$$$ anisotropy into TEdDI (STEdDI).
Approach: We simulate $$$R_2$$$ of PGSE signal in magnetized cylinders using Monte-Carlo, and fit TEdDI and STEdDI on ex vivo mouse multi-echo dMRI data acquired at 16.4T.
Results: $$$R_2$$$ anisotropy outside axons are non-axially-symmetric, depends on B0 direction, gradient direction and b-value. Residuals are significantly lower with STEdDI in dMRI data.
Impact: Interplay between microscopic magnetic fields and diffusion weighting affects $$$R_2$$$ in extra-axonal space. Incorporating $$$R_2$$$ anisotropy$$$\,$$$into modeling lowered the residuals and may allow rotation-free estimation of $$$R_2$$$ anisotropy, which could be useful to gain a deeper insight into brain microstructure.
Introduction
Estimating all the parameters of the Standard Model (SM) of white matter (WM) is ill-posed1–4. It has been shown that incorporating isotropic transverse relaxation ($$$R_2$$$) to the SM5–7 signal kernel helps estimating compartmental relaxation rates but with little improvement on SM parameters. In addition, several studies5,8–11 have found $$$R_2\,$$$to be anisotropic presumably due to mesoscopic susceptibility effects, and when applying diffusion gradients, mesoscopic susceptibility effects can bias the apparent diffusivity from extra-axonal water5,8,9,12–18. Our main question here is: How is apparent R2 affected by diffusion weighting and magnetic susceptibility in WM, and can we incorporate it into SM? We investigate $$$R_2\,$$$anisotropy of a multi-echo PGSE signal with Monte-Carlo (MC) simulations using a digital phantom made of parallel cylinders. We acquire multi-echo dMRI data of an ex-vivo mouse brain and fit our proposed model STEdDI – a minimal extension of the SM model TEdDI6 where S denotes adding susceptibility-related orientation dependent extra-axonal $$$R_2$$$. The effect of $$$R_2\,$$$anisotropy in the SM kernel was previously analyzed by Tax et. al5, but to our knowledge, fitting the parameters with R2 anisotropy added to the SM kernel has not been pursued. We also fit TEdDI and compare their Bayesian Information Criteria (BIC)19.Methods
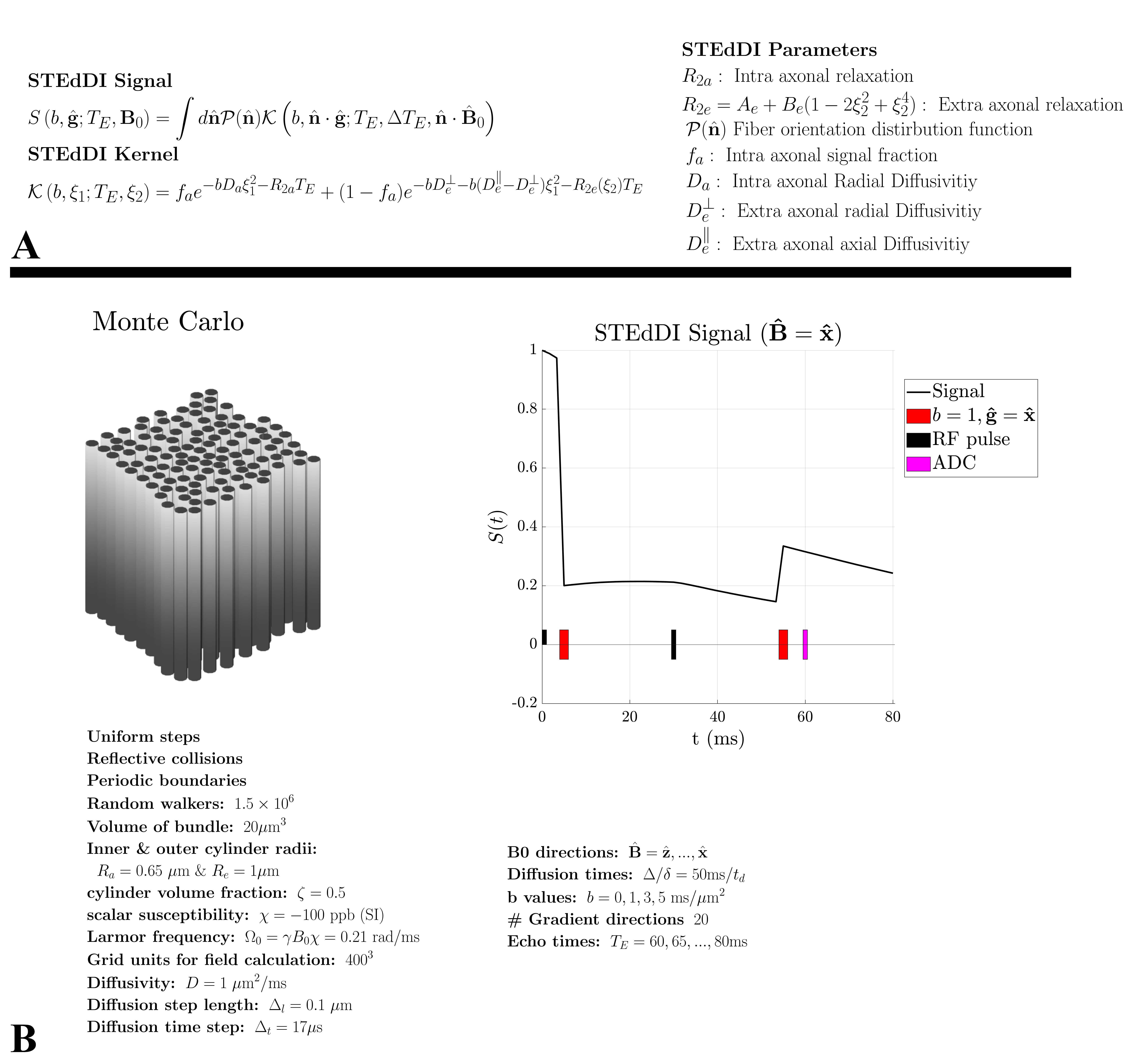
TheoryWe propose STEdDI, a minimal extension of TEdDI, which models $$$R_2\,$$$for extra-axonal water as $$$R_{2e}(\theta_\mathrm{\hat{B}})=A+B\sin^4(\theta_\mathrm{\hat{B}})$$$, valid for long diffusion times. Here $$$\theta_\mathrm{\hat{B}}\,$$$is the angle between cylinder axis $$$\mathrm{\hat{n}}\,$$$and the scanner $$$\mathrm{{B}_0}=B_0\mathrm{\hat{B}}$$$. This $$$R_{2e}\,$$$anisotropy stems from the microscopic fields generated outside magnetized cylinders (axons) – irrespective of susceptibility anisotropy20,21. An overview of STEdDI is shown in figure 1A.
Monte-Carlo simulations
We performed MC random walk simulations to investigate the apparent extra-axonal $$$R_2$$$ originating from the interplay between diffusion gradients $$$\mathrm{{g}}$$$ and the induced field $$$\mathrm{\Delta B}$$$, for a bundle of randomly packed parallel cylinders. The details are listed in figure 1B.
Ex vivo mouse brain
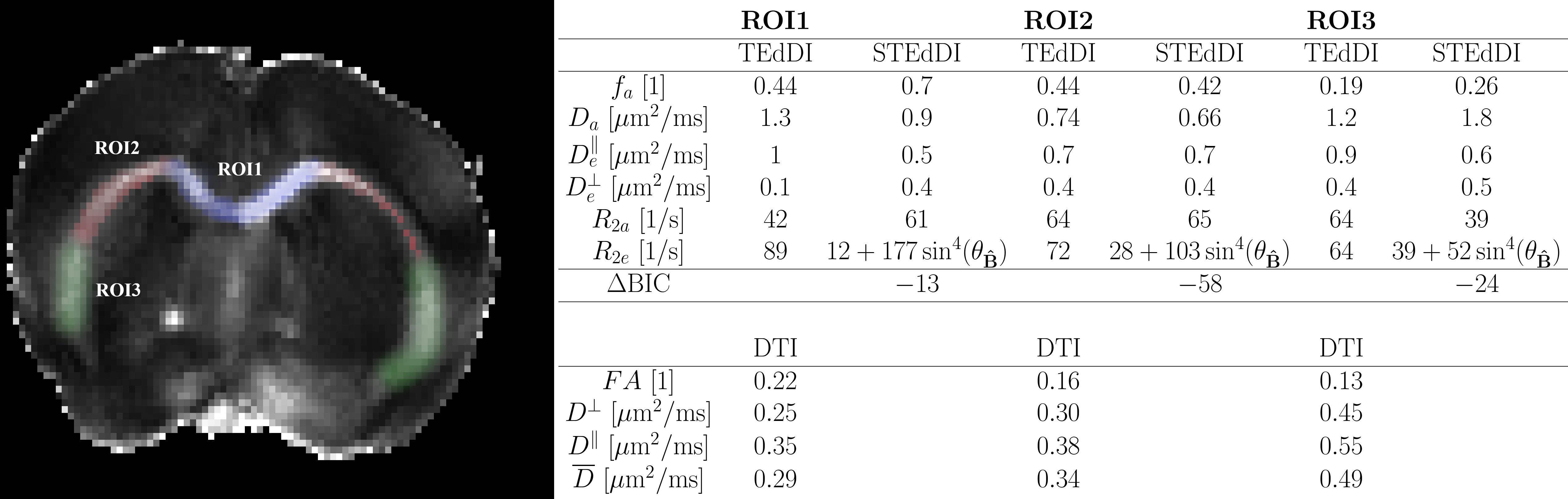
We acquired multi-echo dMRI data of a mouse brain to fit STEdDI and TEdDI (see acquisition details in figure 5 caption). We fitted the signal from three manually segmented ROIs of Corpus Callosum (see figure 5). We then compared the models by computing their difference in BIC $$$(\mathrm{\Delta BIC})$$$18.
Results
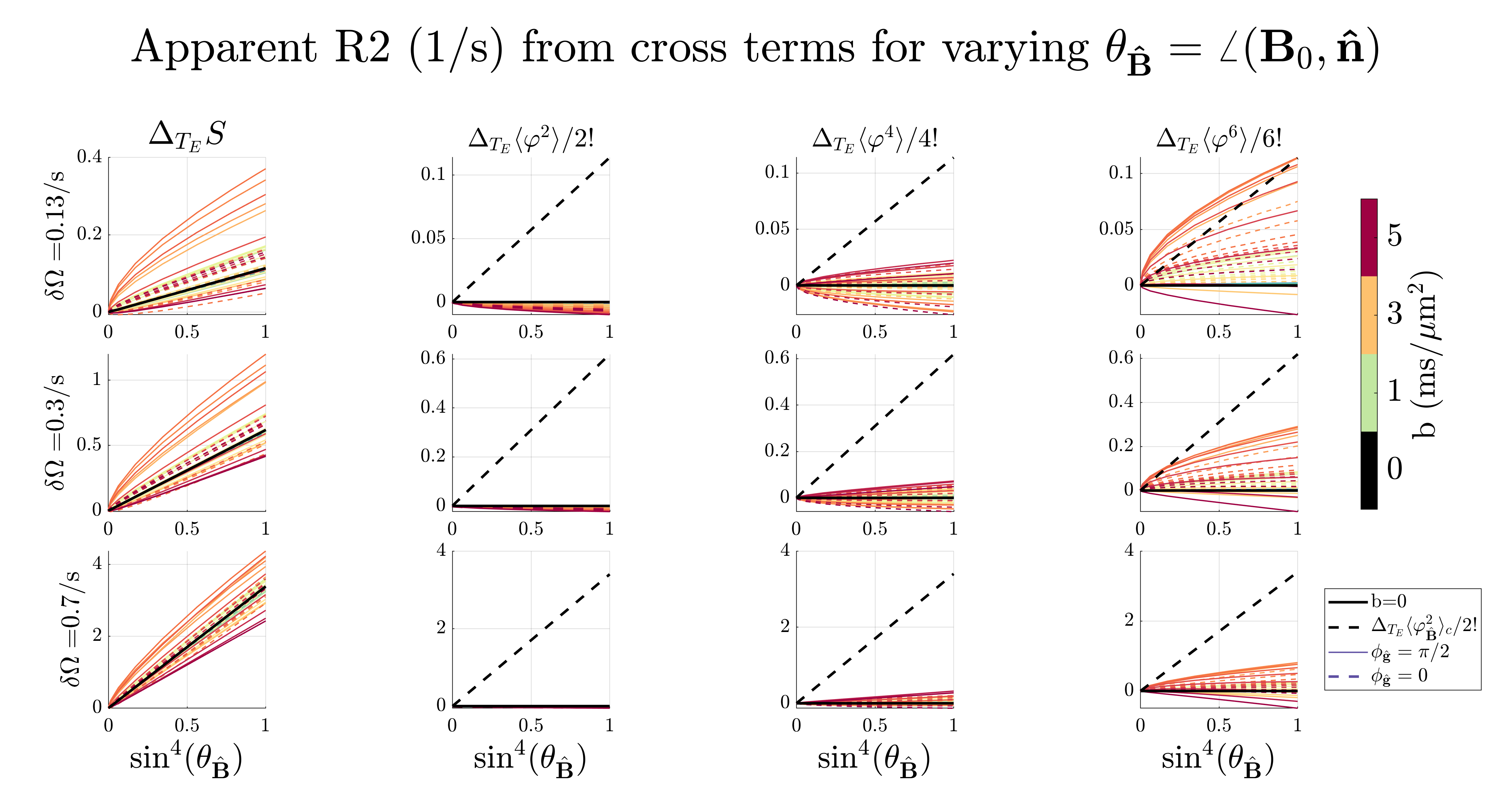
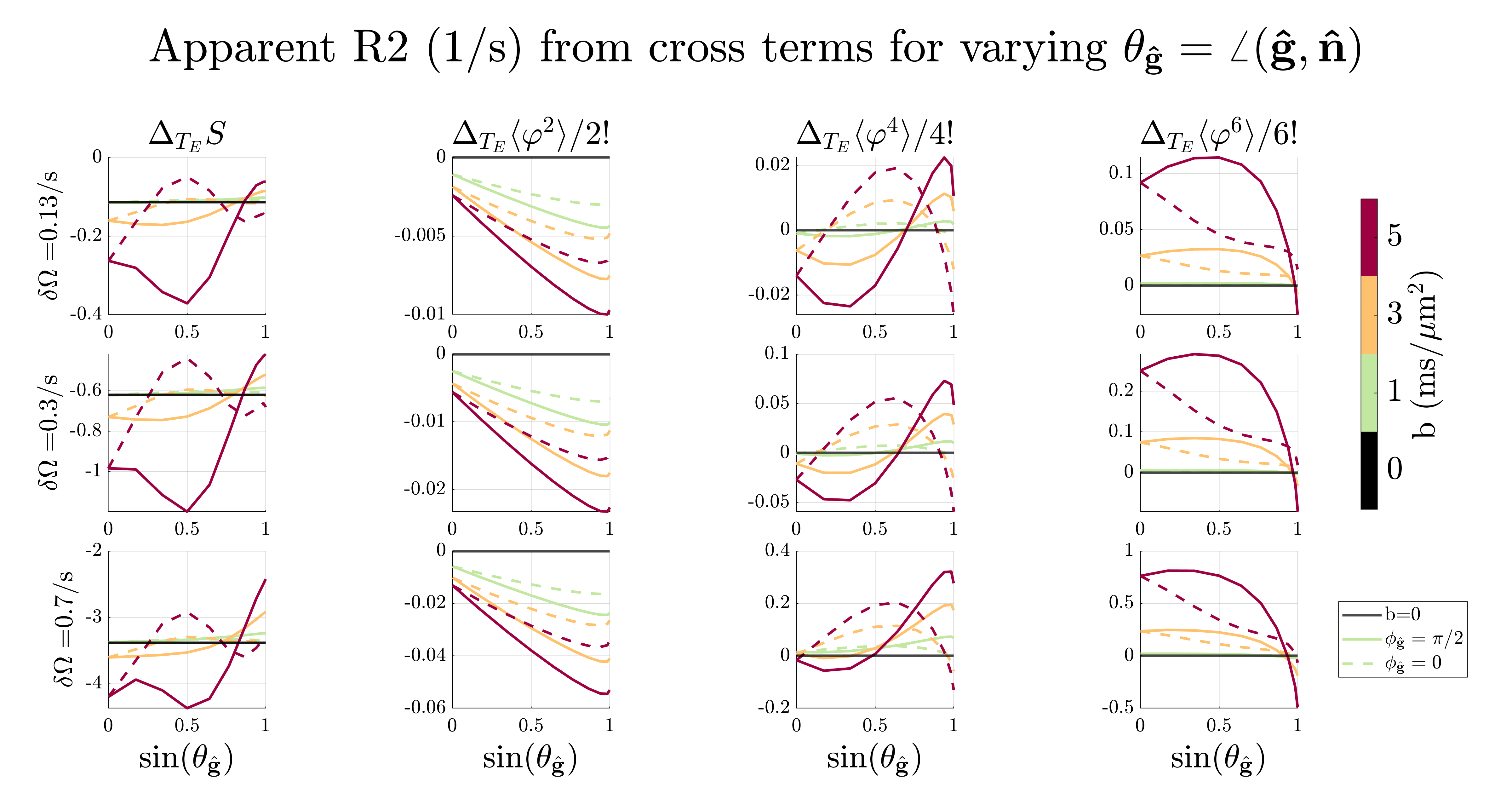
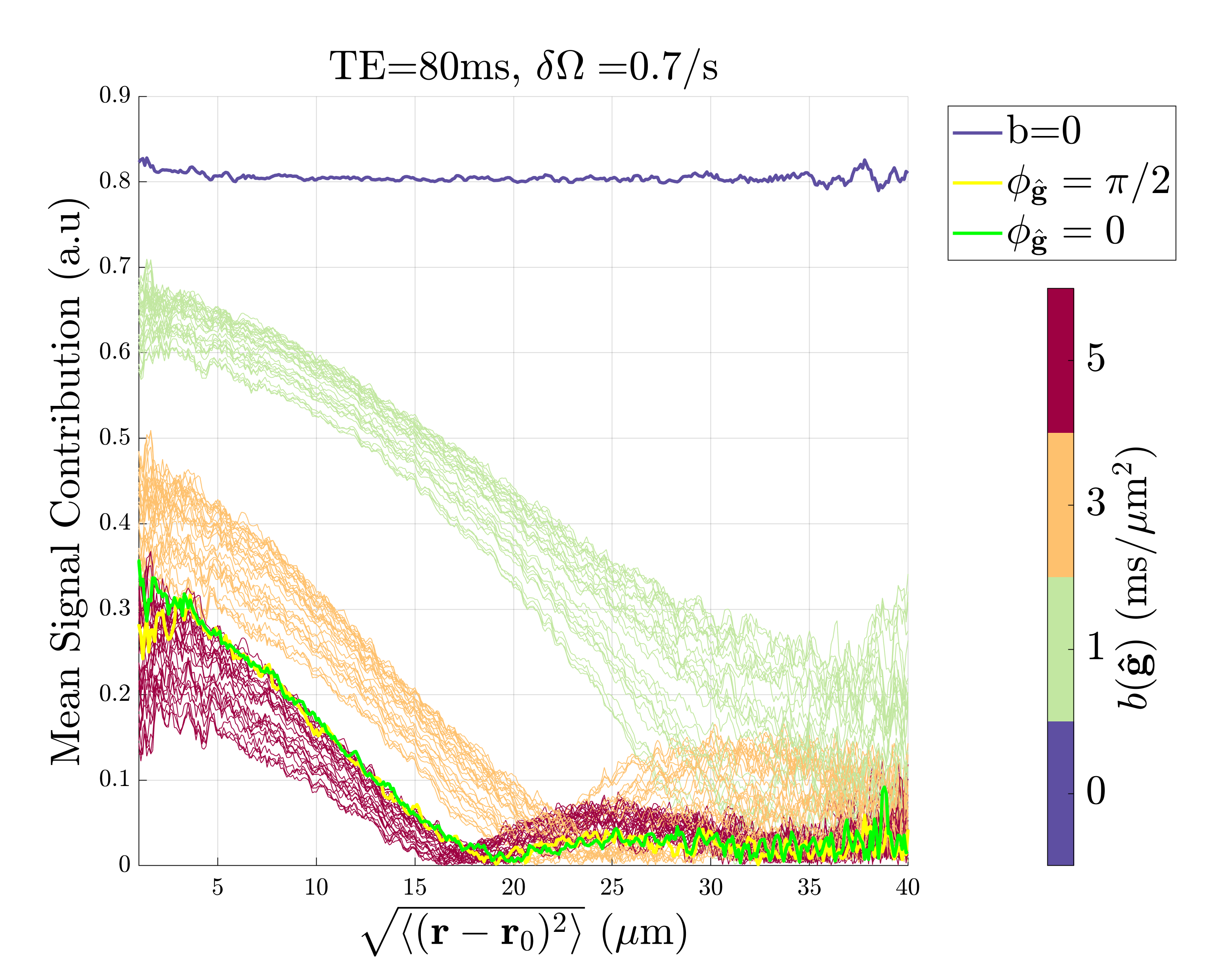
MC simulationsFigure 2-3 shows the apparent $$$R_2\,$$$from the whole extra-axonal signal21-23 and the contribution from the n’th cumulants $$$\langle\varphi^n\rangle\equiv \langle(\varphi_\mathrm{\hat{B}}+\varphi_\mathrm{\hat{g}})^n\rangle_c - \langle\varphi^n_\mathrm{\hat{B}}\rangle_c-\langle\varphi^n_\mathrm{\hat{g}}\rangle_c$$$ due to $$$\mathrm{{g}}$$$ and $$$\mathrm{\Delta B}$$$. It is clear that the phase from diffusion gradients $$$\varphi_\mathrm{\hat{g}}$$$ interacts with the phase from internal fields $$$\varphi_\mathrm{\hat{B}}$$$, making $$$R_2\,$$$deviate from $$$\sin^4(\theta_\mathrm{\hat{B}})$$$. The 6th cumulant cross-term $$$\langle\varphi^6\rangle\,$$$produced the highest relaxation, but as the internal field variance $$$\delta\Omega=\gamma B_0\sqrt{\mathrm{\hat{B}}^\mathrm{T}\cdot\mathrm{\Delta{B}}}\,$$$increased, its contribution became negligible compared to $$$\langle\varphi^2_\mathrm{\hat{B}}\rangle_c$$$. Figure 4 shows the spins' signal contribution versus root-mean-square displacement. The total signal relaxation vs. $$$T_E\,$$$is also weighted differently depending on $$$\mathrm{{g}}$$$, which in turn modulated the signal in a $$$\mathrm{\Delta B}$$$-dependent way. The non-axially symmetric contribution comes from spins near the cylinders (see green and yellow curve in figure 3 and figure 4), where $$$\mathrm{g}\,$$$and $$$\mathrm{\Delta B}\,$$$can interfere constructively or destructively, respectively.
Ex vivo mouse brain
Figure 5 shows the estimated parameters in the CC-ROIs by fitting TEDdi and STEdDI, respectively. Adding $$$R_2\,$$$anisotropy had a substantial impact on parameter values. We found $$$(\mathrm{\Delta BIC})<-6\,$$$in all ROIs thus favoring STEdDI over TEdDI25.
Discussion
Our simulations demonstrate how $$$R_2\,$$$of a multi-echo PGSE signal depend on the interaction between the induced magnetic field from cylinders and diffusion gradients in the limit of long diffusion times, especially when the applied gradients were strong compared to $$$B_0$$$. These correlations may therefore be important at 3T and/or high gradient scanners like the Connectome. We plan to investigate this analytically in the future. Here we demonstrate that it may be feasible to include $$$R_2\,$$$anisotropy in TEdDI and estimate it without having to rotate the sample. This is because orientation dispersion also makes $$$R_2\,$$$vary depending on the applied diffusion gradient26. We find $$$R_2\,$$$anisotropy to be in agreement with Tax et al5 when scaling by the field-strength ratio$$$\,{(16.4\text{T}/3\text{T})^2}$$$. Planar encoding27,28 may increase the sensitivity to $$$R_2\,$$$anisotropy, as it allows one to probe the signal from axons along a single axis. However, planar encoding comes at the expense of lower SNR29, and potentially stronger correlations with internal magnetic fields. This will also be investigated in the future.Conclusion
The apparent$$$\,R_2\,$$$of the PGSE signal from water outside long magnetized cylinders is anisotropic and depends on experimental settings like diffusion weighting and $$$B_0$$$. Incorporating $$$R_2\,$$$anisotropy into the Standard Model of diffusion in white mater lowered the fit residuals without adding too much model complexity and was estimated in an ex-vivo mouse brain without acquiring data from multiple sample orientations.Acknowledgements
The study is funded by the Independent Research Fund Denmark (Grant 10.46540/3103-00144B).References
1. Novikov DS, Fieremans E, Jespersen SN, Kiselev VG. Quantifying brain microstructure with diffusion MRI: Theory and parameter estimation. NMR Biomed. 2019;32(4):e3998. doi:10.1002/NBM.3998
2. Novikov DS, Veraart J, Jelescu IO, Fieremans E. Rotationally-invariant mapping of scalar and orientational metrics of neuronal microstructure with diffusion MRI. Neuroimage. 2018;174:518-538. doi:10.1016/J.NEUROIMAGE.2018.03.006
3. Jelescu IO, Veraart J, Fieremans E, Novikov DS. Degeneracy in model parameter estimation for multi-compartmental diffusion in neuronal tissue. NMR Biomed. 2016;29(1):33-47. doi:10.1002/NBM.3450 4. Coelho S, Pozo JM, Jespersen SN, Jones DK, Frangi AF. Resolving degeneracy in diffusion MRI biophysical model parameter estimation using double diffusion encoding. Magn Reson Med. 2019;82(1):395-410. doi:10.1002/MRM.27714
5. Tax CMW, Kleban E, Chamberland M, Baraković M, Rudrapatna U, Jones DK. Measuring compartmental T2-orientational dependence in human brain white matter using a tiltable RF coil and diffusion-T2 correlation MRI. Neuroimage. 2021;236. doi:10.1016/J.NEUROIMAGE.2021.117967
6. Veraart J, Novikov DS, Fieremans E. TE dependent Diffusion Imaging (TEdDI) distinguishes between compartmental T2 relaxation times. Neuroimage. 2018;182:360-369. doi:10.1016/j.neuroimage.2017.09.030
7. Lampinen B, Szczepankiewicz F, Mårtensson J, et al. Towards unconstrained compartment modeling in white matter using diffusion-relaxation MRI with tensor-valued diffusion encoding. Magn Reson Med. 2020;84(3):1605-1623. doi:10.1002/MRM.28216
8. Knight MJ, Dillon S, Jarutyte L, Kauppinen RA. Magnetic Resonance Relaxation Anisotropy: Physical Principles and Uses in Microstructure Imaging. Biophys J. 2017;112(7):1517-1528. doi:10.1016/j.bpj.2017.02.026
9. Knight MJ, Kauppinen RA. Diffusion-mediated nuclear spin phase decoherence in cylindrically porous materials. J Magn Reson. 2016;269:1-12. doi:10.1016/J.JMR.2016.05.007
10. Veronica P Dell’Acqua, Tax CMW, Malwina Molendowska, Jones DK, Kleban E, Greg D Parker. Measuring compartmental T2 and T2* orientation dependence in white matter. In: ISMRM Workshop on Diffusion MRI: From Research to Clinic. ; 2022.
11. Gil R, Khabipova D, Zwiers M, Hilbert T, Kober T, Marques JP. An in vivo study of the orientation-dependent and independent components of transverse relaxation rates in white matter. NMR Biomed. 2016;29(12):1780-1790. doi:10.1002/NBM.3616
12. Kiselev VG. Effect of magnetic field gradients induced by microvasculature on NMR measurements of molecular self-diffusion in biological tissues. J Magn Reson. 2004;170(2):228-235. doi:10.1016/J.JMR.2004.07.004
13. Does MD, Zhong J, Gore JC. In Vivo Measurement of ADC Change Due to Intravascular Susceptibility Variation. doi:10.1002/(SICI)1522-2594(199902)41:2
14. Zhong J, Kennan RP, Gore JC. Effects of susceptibility variations on NMR measurements of diffusion. J Magn Reson. 1991;95(2):267-280. doi:10.1016/0022-2364(91)90217-H
15. Novikov DS, Reisert M, Kiselev VG. Effects of mesoscopic susceptibility and transverse relaxation on diffusion NMR. J Magn Reson. 2018;293:134-144. doi:10.1016/J.JMR.2018.06.007
16. Perez LAP, Alvarez GA. Cumulant expansion framework for internal gradient distributions tensors. Published online April 4, 2023. Accessed July 5, 2023. http://arxiv.org/abs/2304.02065
17. Álvarez GA, Shemesh N, Frydman L. Internal gradient distributions: A susceptibility-derived tensor delivering morphologies by magnetic resonance. Sci Rep. 2017;7(1):1-10. doi:10.1038/s41598-017-03277-9
18. Winther S, Lundell H, Rafael-Patiño J, Andersson M, Thiran JPJP, Dyrby TB. Susceptibility-induced internal gradients reveal axon morphology and cause anisotropic effects in the dMRI signal. bioRxiv. Published online May 2, 2023:2023.05.01.538981. doi:10.1101/2023.05.01.538981
19. Schwarz G. Estimating the Dimension of a Model. https://doi.org/101214/aos/1176344136. 1978;6(2):461-464. doi:10.1214/AOS/1176344136
20. Sukstanskii AL, Yablonskiy DA. On the role of neuronal magnetic susceptibility and structure symmetry on gradient echo MR signal formation. Magn Reson Med. 2014;71(1):345-353. doi:10.1002/mrm.24629
21. Wharton S, Bowtell R. Fiber orientation-dependent white matter contrast in gradient echo MRI. Proc Natl Acad Sci U S A. 2012;109(45):18559-18564. doi:10.1073/pnas.1211075109
22. Sukstanskii AL, Yablonskiy DA. Gaussian approximation in the theory of MR signal formation in the presence of structure-specific magnetic field inhomogeneities. Effects of impermeable susceptibility inclusions. J Magn Reson. 2004;167(1):56-67. doi:10.1016/J.JMR.2003.11.006
23. Kiselev VG, Posse S. Analytical Model of Susceptibility-Induced MR Signal Dephasing: Effect of Diffusion in a Microvascular Network. Magn Reson Med. 1999;41:499-509. doi:10.1002/(SICI)1522-2594(199903)41:3
24. Frøhlich AF, Østergaard L, Kiselev VG. Theory of susceptibility-induced transverse relaxation in the capillary network in the diffusion narrowing regime. Magn Reson Med. 2005;53(3):564-573. doi:10.1002/MRM.20394
25. Kass RE, Raftery AE. Bayes factors. J Am Stat Assoc. 1995;90(430):773-795. doi:10.1080/01621459.1995.10476572
26. Sandgaard AD, Kiselev VG, Shemesh N, Jespersen SN. Incorporating susceptibility effects into the standard model of diffusion in white matter. In: Joint Annual Meeting ISMRM-ESMRMB & ISMRT 32st Annual Meeting. ; 2023:0975.
27. Dhital B, Reisert M, Kellner E, Kiselev VG. Intra-axonal diffusivity in brain white matter. Neuroimage. 2019;189:543-550. doi:10.1016/J.NEUROIMAGE.2019.01.015
28. Westin CF, Szczepankiewicz F, Pasternak O, et al. Measurement tensors in diffusion MRI: Generalizing the concept of diffusion encoding. Lect Notes Comput Sci (including Subser Lect Notes Artif Intell Lect Notes Bioinformatics). 2014;8675 LNCS(PART 3):209-216. doi:10.1007/978-3-319-10443-0_27/COVER
29. Coelho S, Baete SH, Lemberskiy G, et al. Reproducibility of the Standard Model of diffusion in white matter on clinical MRI systems. Neuroimage. 2022;257:119290. doi:10.1016/J.NEUROIMAGE.2022.119290
30. Veraart J, Novikov DS, Christiaens D, Ades-aron B, Sijbers J, Fieremans E. Denoising of diffusion MRI using random matrix theory. Neuroimage. 2016;142:394-406. doi:10.1016/j.neuroimage.2016.08.016 31. Olesen JL, Ianus A, Østergaard L, Shemesh N, Jespersen SN. Tensor denoising of high-dimensional MRI data. Published online March 30, 2022. doi:10.48550/arxiv.2203.16078
32. Kellner E, Dhital B, Kiselev VG, Reisert M. Gibbs-ringing artifact removal based on local subvoxel-shifts. Magn Reson Med. 2016;76(5):1574-1581. doi:10.1002/mrm.26054
Figures