3456
Diffusion in dendritic spines: impact on permeative exchange estimation with time-dependent diffusion-weighted MRI1Cardiff University Brain Research Imaging Centre (CUBRIC), School of Psychology, Cardiff University, Cardiff, United Kingdom, 2School of Computer Science and Informatics, Cardiff University, Cardiff, United Kingdom
Synopsis
Keywords: Microstructure, Microstructure, diffusion, brain, exchange, gray matter, water, microstructure, simulation, spines, dendrites
Motivation: Diffusion exchange models NEXI and SMEX assumes permeative exchange between intra- and extra-neurite compartments. Here, we hypothesize fine microstructures, like spines in dendritic segments, without permeable membranes can mimic permeative exchange.
Goal(s): We aim to emphasize the significance of taking diffusion-mediated exchange into account when interpreting model-based estimates of exchange in intricate microstructures like the gray matter of the brain.
Approach: Monte-Carlo water diffusion simulations in spiny dendritic branches, also featuring undulations and beading
Results: Our results question the way we interpret dMRI exchange estimates, emphasizing the need to exercise caution when inferring these estimates solely as indicators of membrane permeability.
Impact: Diffusion exchange models assumes permeative exchange between intra- and extra-neurite compartments. We hypothesize fine microstructures without permeable membranes can mimic permeative exchange. We aim to emphasize considering the impact of intricate microstructures in gray matter when interpreting model-based exchange estimates
Introduction
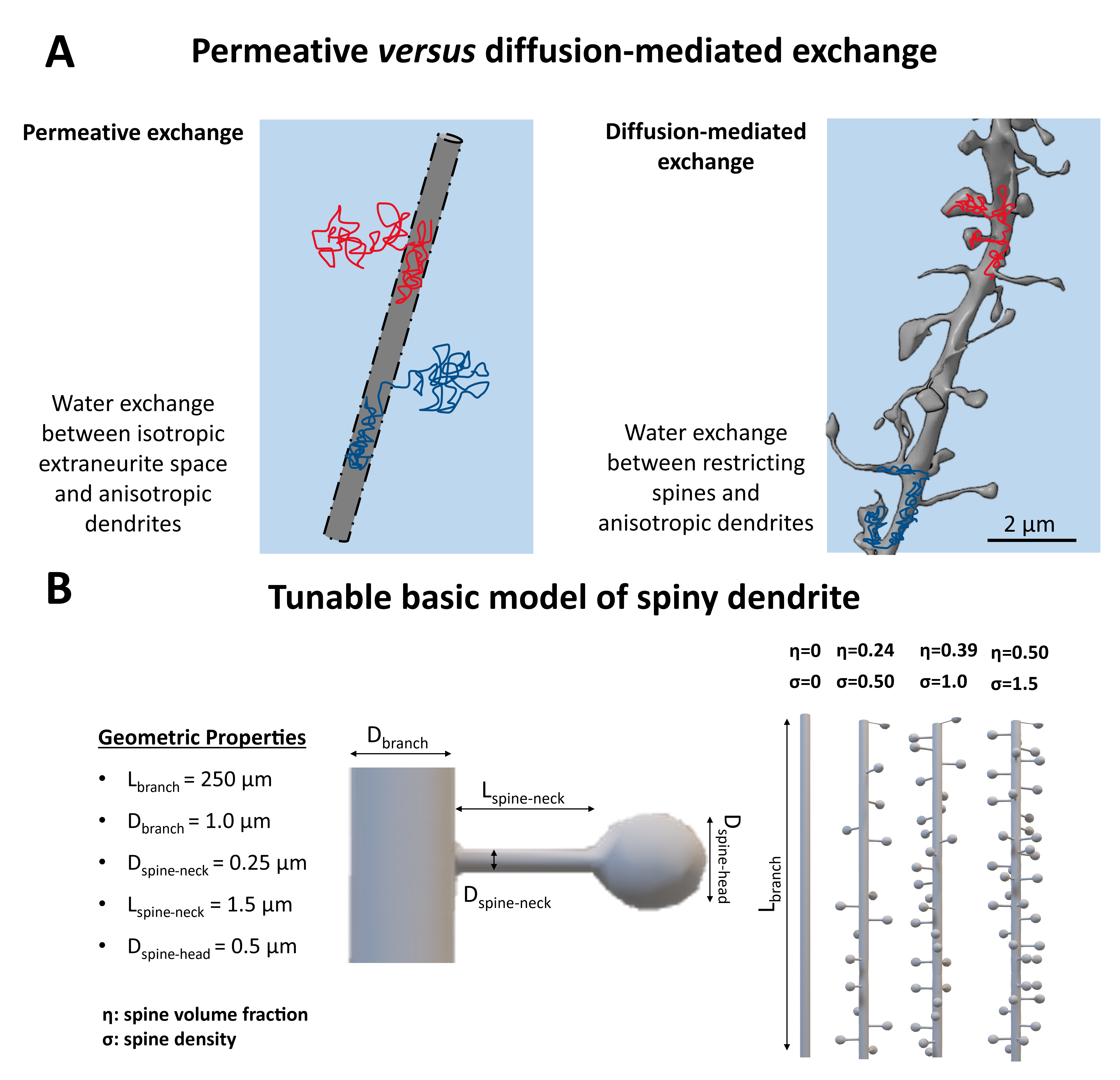
Time-dependent diffusion-weighted MRI (dMRI) can probe exchange in complex biological tissues1–4. While it is expected that different exchange mechanisms contribute to the time-dependent signal5,6, measurements are often primarily interpreted in terms of transcytolemmal water exchange. The Neurite Exchange Imaging (NEXI)3 and the Standard Model with Exchange (SMEX)2 are two recent examples of model-based approaches to estimate exchange in brain tissue. Both NEXI and SMEX assume that the only exchange mechanism leading to the measured time-dependent dMRI signal is permeative exchange between intra and extra-neurite compartments.This work aims to underscore the importance of considering diffusion-mediated exchange when interpreting model-based estimates of exchange in complex microstructures such as the brain Gray Matter (GM). We hypothesize that water diffusing within spiny dendrites in the GM, without crossing the cell membrane, can originate a time-dependent signature which is indistinguishable from permeative exchange (Fig.1A); and use Monte-Carlo simulations with a basic model of spiny dendrite (Fig.1B) to validate it and evaluate the impact of different spine densities on NEXI/SMEX estimates.
Methods
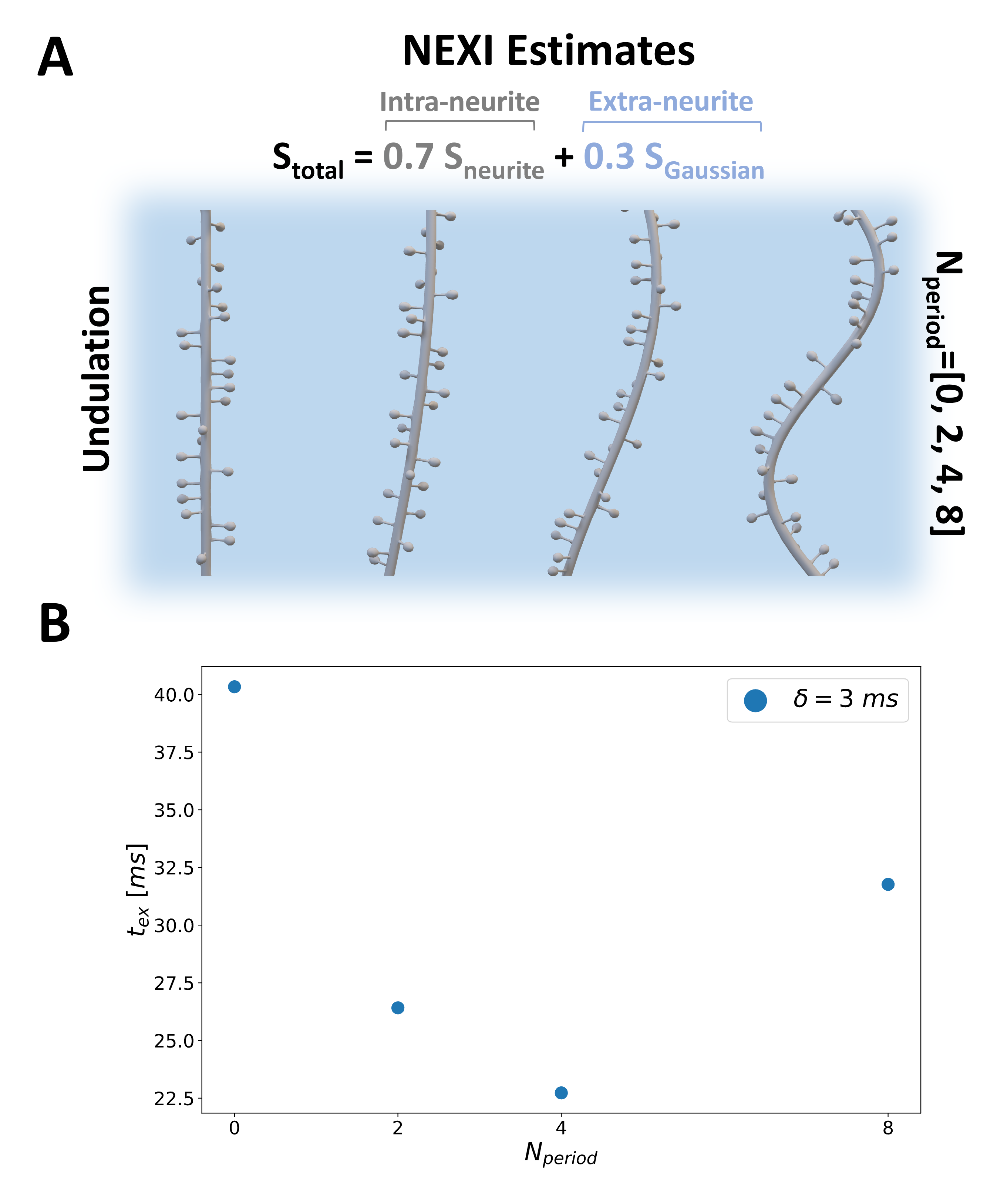
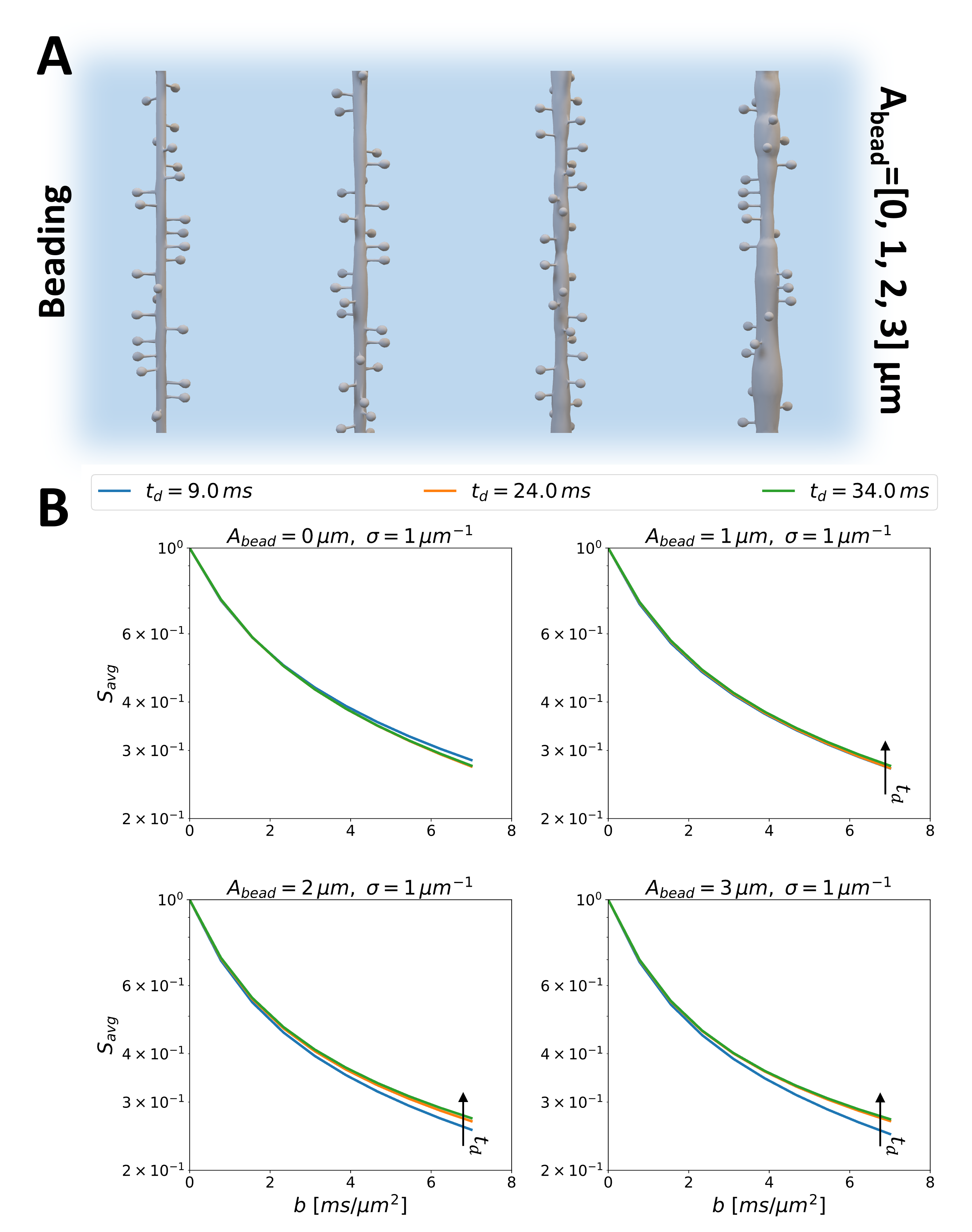
Spiny Dendritic MeshesSkeletons of spiny dendritic branches for two sets of substrates were built on MathWorks MATLAB 2022a7 involving functions from the Trees-Toolbox8 and then surface meshed using Python Blender API v2.799. Set I: ten spiny branches ($$$\sigma$$$=[0, 2.25] μm-1) for investigating the exchange effect of diffusion in spines without undulation and/or beading. Set II: 16 spiny branches ($$$\sigma$$$=1μm-1), featuring combinations of 4 undulation periods ($$$N_{period}$$$=[0, 2, 4, 8]) and 4 beading amplitudes ($$$A_{bead}$$$=[0, 1, 2, 3] μm). Other details of the spine morphology can be found in Fig.1A. Notably, any of these features can be changed arbitrarily, here, we investigated realistic spine densities and realistic to extreme undulations and beading in dendritic branches.
Diffusion Simulations & Data Analysis
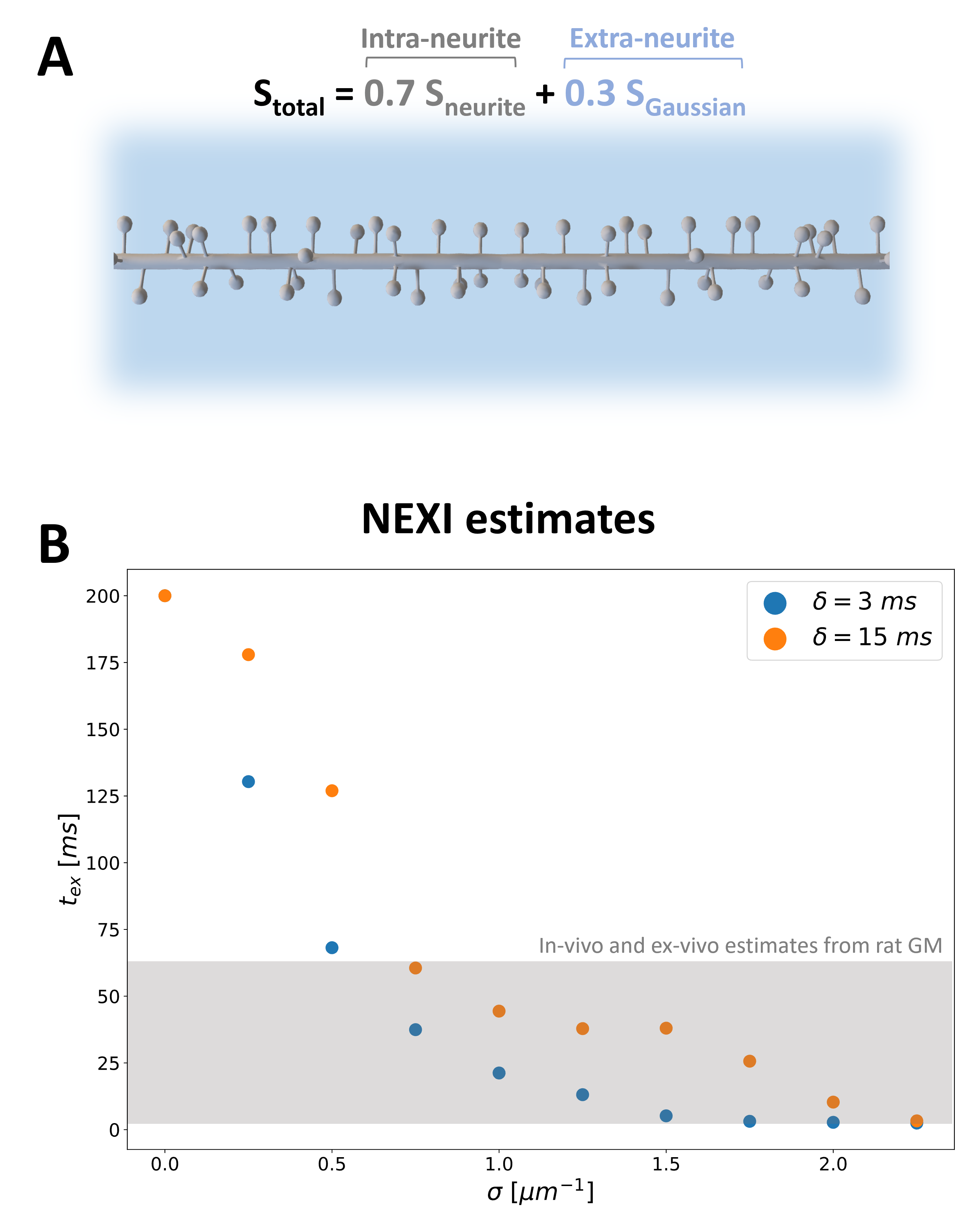
DisimPy10 was employed in all simulations. The number of spins and time steps were determined by the Monte-Carlo convergence11,12:106 and 2000, respectively. Five different pulsed gradient schemes, combinations of three gradient separations ($$$\Delta$$$=[10, 25, 35] ms) and two gradient durations ($$$\delta$$$=[3, 15] ms) were used with 128 directions and diffusion-weighting b up to 7ms/μm2. Periodic boundary conditions were used for intra-branch diffusion simulations with diffusivities 2μm2/ms (typical value for intra-neurite water2,13). For a fair comparison with NEXI/SMEX biophysical modelling, we simulated the total signal arising from a dMRI voxel as the weighted sum of intra and extra-neurite signals, $$$S_{neurite}$$$ and $$$S_{Gaussian}$$$, respectively: $$$S_{total}=0.7S_{neurite}+0.3S_{Gaussian}$$$. $$$S_{neurite}$$$ is derived from our simulation in spiny dendrites at different spine densities and/or undulations, while $$$S_{Gaussian}$$$ is mono-exponential decay with diffusivity of 1 µm2/ms (Fig.3,4). We assumed 70% intra-neurite signal fraction. $$$S_{total}$$$ was fitted using NEXI model3 to estimate the exchange time ($$$t_{ex}$$$) for all simulated conditions.
Results and Discussion
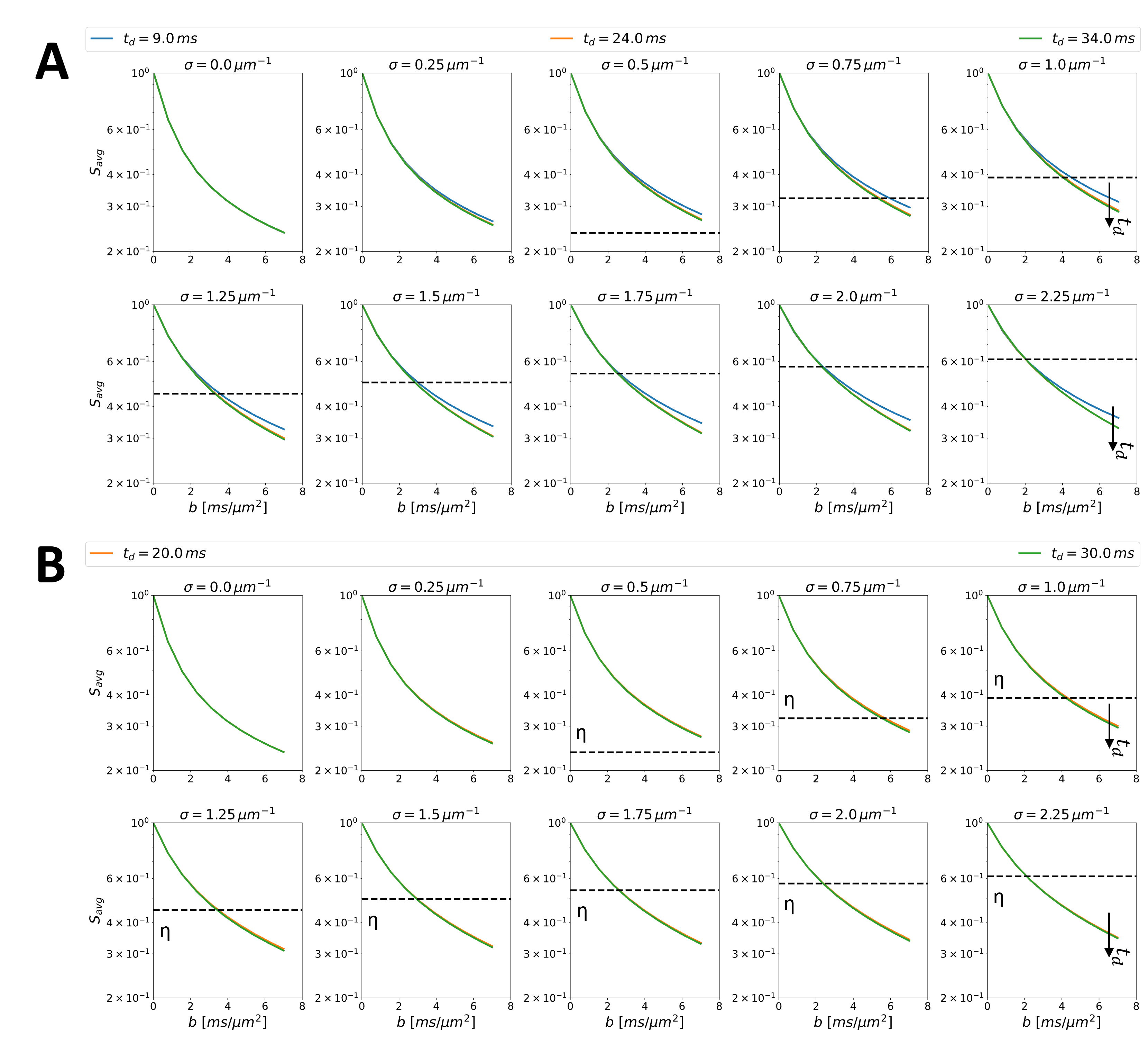
Fig.2 shows the time-dependence of simulated signals at different b-values and for different spine densities. Decreasing signal amplitude with increasing $$$t_d$$$ (at fixed $$$b$$$) is more marked at higher spine densities and mirrors the observed time-dependence in both ex-vivo2 and in-vivo3 measurements in the rat GM. These time-dependencies have been interpreted solely as a consequence of membrane permeability. Fig.2 documents that the presence of spines can also lead to similar observation without any permeation across the dendritic membrane.Furthermore, Fig.3B shows the exchange time $$$t_{ex}$$$ estimated using NEXI when we assume that the total signal in the dMRI voxel comes from a weighted sum of intra and extra-neurite signals, with spiny dendrites and no membrane permeability (Fig.3A). The estimated $$$t_{ex}$$$ values (3-60 ms) for spine densities typically observed in healthy cortical GM (0.5–1.5 µm-1) are in very good agreement with both in-vivo and ex-vivo estimates from rat GM2,3 and human14.
Finally, we acknowledge that the simulated spiny dendrites are oversimplified. Other microstructural features, such as undulations and beading, could also confound the interpretation of exchange mechanisms. Indeed, adding undulations leads to shorter exchange times when estimated with NEXI for realistic undulation periods 0-4 (Fig.4). In contrast, Fig.5 shows that beading leads to a different time-dependence of the direction-averaged signal, indicating more restriction to the diffusion, as previously reported15.
Future work will investigate whether it is possible to disentangle permeative from diffusion-mediated exchange and how sensitive and specific our measurements can be to spine-induced diffusion-mediated exchange (e.g. with respect to undulations and beading).
Conclusion
The time-dependent signal from diffusion within impermeable spiny dendrites is indistinguishable from permeative exchange. Our findings can contribute to a more insightful interpretation of exchange estimates using dMRI, underscoring the importance of cautioning when inferring exchange estimates as indicators of membrane permeability only.Acknowledgements
- This work, KS and MP are supported by UKRI Future Leaders Fellowship (MR/T020296/2).
References
1. Kärger J. Der Einfluß der Zweibereichdiffusion auf die Spinechodämpfung unter Berücksichtigung der Relaxation bei Messungen mit der Methode der gepulsten Feldgradienten. Ann Phys. 1971;482(1):107-109. doi:10.1002/ANDP.19714820113
2. Olesen JL, Østergaard L, Shemesh N, Jespersen SN. Diffusion time dependence, power-law scaling, and exchange in gray matter. Neuroimage. 2022;251:118976. doi:10.1016/J.NEUROIMAGE.2022.118976
3. Jelescu IO, de Skowronski A, Geffroy F, Palombo M, Novikov DS. Neurite Exchange Imaging (NEXI): A minimal model of diffusion in gray matter with inter-compartment water exchange. Neuroimage. 2022;256:119277. doi:10.1016/J.NEUROIMAGE.2022.119277
4. Lasič S, Nilsson M, Lätt J, Ståhlberg F, Topgaard D. Apparent exchange rate mapping with diffusion MRI. Magn Reson Med. 2011;66(2):356-365. doi:10.1002/MRM.22782
5. Khateri M, Reisert M, Sierra A, Tohka J, Kiselev VG. What does FEXI measure? NMR Biomed. 2022;35(12):e4804. doi:10.1002/NBM.4804
6. Bai R, Li Z, Sun C, Hsu YC, Liang H, Basser P. Feasibility of filter-exchange imaging (FEXI) in measuring different exchange processes in human brain. Neuroimage. 2020;219:117039. doi:10.1016/J.NEUROIMAGE.2020.117039
7. The MathWorks Inc. MATLAB version: 9.12.0 (R2022a). Published online 2022.
8. Cuntz H, Forstner F, Borst A, Häusser M. One Rule to Grow Them All: A General Theory of Neuronal Branching and Its Practical Application. PLoS Comput Biol. 2010;6(8):e1000877. doi:10.1371/JOURNAL.PCBI.1000877
9. Blender Online Community. Blender - a 3D modelling and rendering package. Published online 2017.
10. Kerkelä L, Nery F, Hall MG, Clark CA. Disimpy: A massively parallel Monte Carlo simulator for generating diffusion-weighted MRI data in Python. J Open Source Softw. 2020;5(52):2527. doi:10.21105/JOSS.02527
11. Hall MG, Alexander DC. Convergence and Parameter Choice for Monte-Carlo Simulations of Diffusion MRI. IEEE Trans Med Imaging. 2009;28(9):1354-1364. doi:10.1109/TMI.2009.2015756
12. Rafael-Patino J, Romascano D, Ramirez-Manzanares A, Canales-Rodríguez EJ, Girard G, Thiran JP. Robust Monte-Carlo Simulations in Diffusion-MRI: Effect of the Substrate Complexity and Parameter Choice on the Reproducibility of Results. Front Neuroinform. 2020;14:457195. doi:10.3389/FNINF.2020.00008/BIBTEX
13. Palombo M, Alexander DC, Zhang H. A generative model of realistic brain cells with application to numerical simulation of the diffusion-weighted MR signal. Neuroimage. 2019;188:391-402. doi:10.1016/J.NEUROIMAGE.2018.12.025
14. Uhl Q, Pavan T, Molendowska M, Jones DK, Palombo M, Jelescu I. Quantifying human gray matter microstructure using NEXI and 300 mT/m gradients. Published online July 18, 2023. Accessed November 8, 2023. https://arxiv.org/abs/2307.09492v1
15. Budde MD, Frank JA. Neurite beading is sufficient to decrease
the apparent diffusion coefficient after ischemic stroke. Proc Natl Acad Sci
U S A. 2010;107(32):14472-14477.
doi:10.1073/PNAS.1004841107/SUPPL_FILE/PNAS.201004841SI.PDF
Figures