3428
Frequency-selective brain state regulation with optogenetic stimulation of central thalamus in unconscious rats1Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital, Charlestown, MA, United States
Synopsis
Keywords: Task/Intervention Based fMRI, Translational Studies, high-field MRI, multimodal, traumatic brain injury
Motivation: There is a lack of effective treatments and understanding of the mechanisms behind disorders of consciousness, including coma.
Goal(s): Explore the mechanisms behind thalamus-mediated subcortical reactivation to promote effective therapies for disorders of consciousness.
Approach: Study brain changes induced by optogenetic stimulation of the central thalamus (CThal) at different frequencies in unconscious rats with fMRI and electrophysiology.
Results: CThal stimulation can trigger brain state changes and electrophysiologically activate the cingulate cortex and other areas of the cortex similar to a more conscious state. BOLD signal activation in the cortex seems to be dependent on the CThal stimulation frequency.
Impact: This study provides useful insights into frequency-dependent central thalamus-mediated brain state changes and offers opportunities to investigate the potential of CThal stimulation in promoting subcortical reactivation and acute coma recovery in a rodent model.
Introduction
Subcortical reactivation, which may be mediated by the central thalamus (CThal), is crucial in acute coma recovery1. However, the detailed mechanisms are still unknown. Here we explore the changes in brain dynamics induced by optogenetic stimulation of CThal at different frequencies with fMRI and electrophysiology.Methods
Long-Evans rats (3.5 weeks old) were injected with two viral vectors to introduce channelrhodopsin in CaMKII+ cells in CThal for optogenetics and for glutamate sensing in the Cingulate Cortex (Cg1). After 3 weeks, the animals were divided into two groups: MRI (n=13) and electrophysiology (n=3). The rats were anesthetized (2% isofluorane) and optogenetically stimulated at CThal while glutamate was recorded in Cg1 with optical fibers inserted. BOLD signal was acquired from the MRI group and Local Field Potentials (LFP) were recorded in Cg1 and the forepaw somatosensory cortex (S1FL) in the electrophysiology group. CThal stimulation was performed at 3, 10 and 40 Hz. Control recordings were also included: without stimulation and while transitioning from deeply anesthetized (2% isofluorane) to a less anesthetized state (0.25% isofluorane).MRI images were acquired with a 14 T/13 mm horizontal-bore scanner equipped with a 6 cm RRI gradient set producing 1 T/m. A transceiver surface coil was used to acquire functional images. fMRI scans with block design were performed using multi-slice 2D Echo planar imaging (EPI) sequence: TR 2 s, TE 5.5 ms, FOV 1.92 x 1.92 x 1.92 cm3, Matrix 64 x 64 x 64 (300 µm isotropic resolution). Optogenetic stimulation block design consisted of an “8s on, 32s off” stimulation paradigm repeated ten times. Three trials were acquired for each condition in each animal.
Results
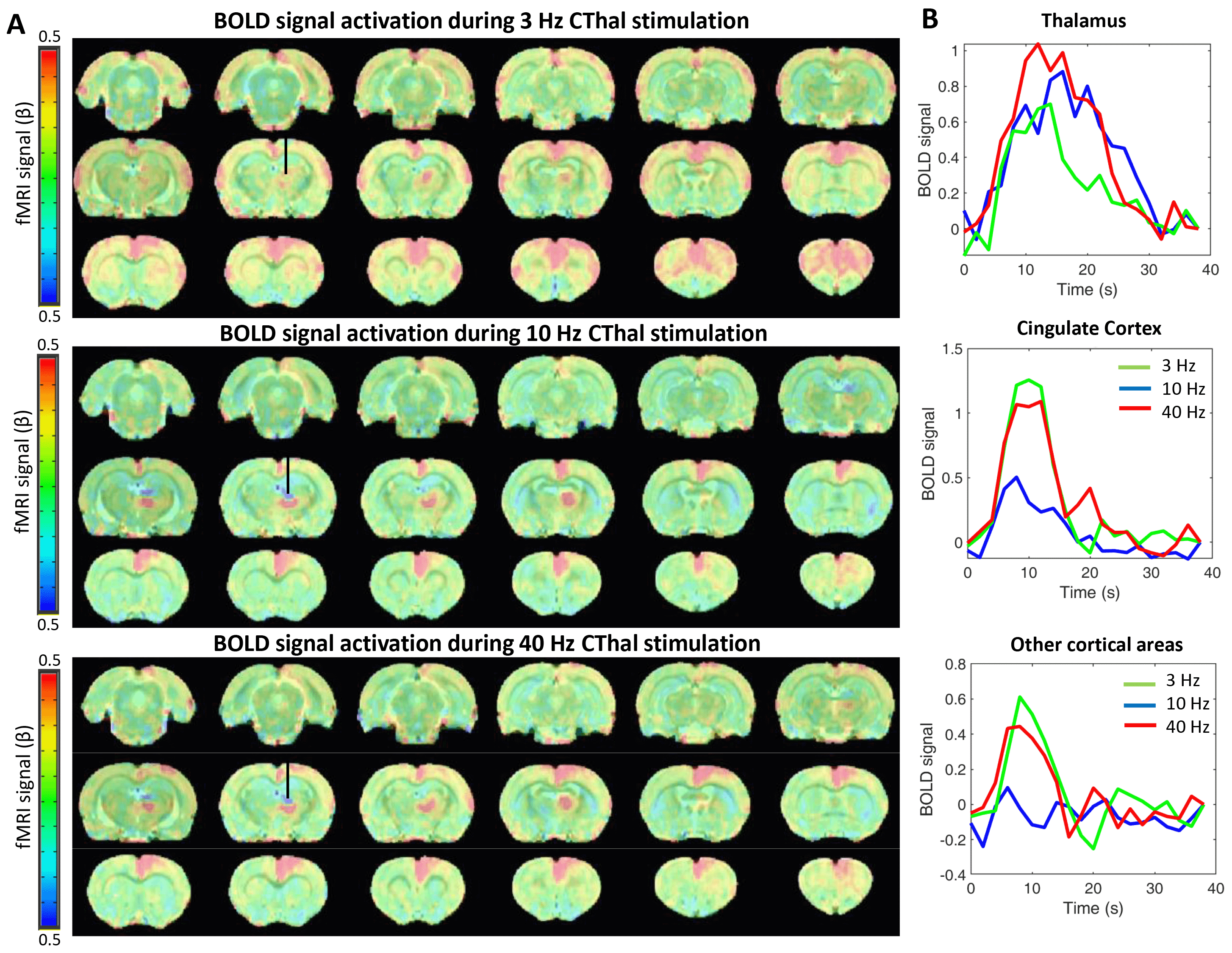
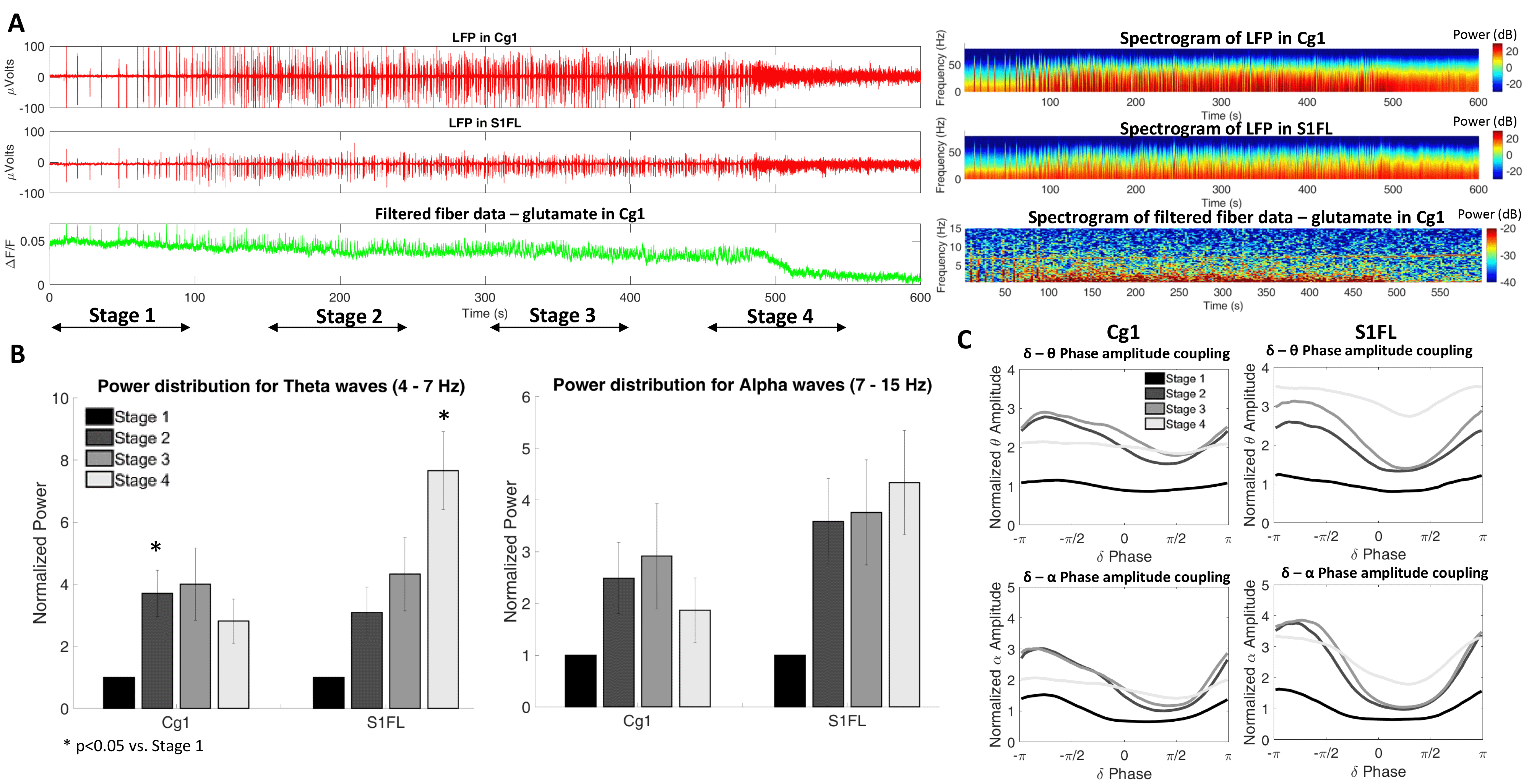
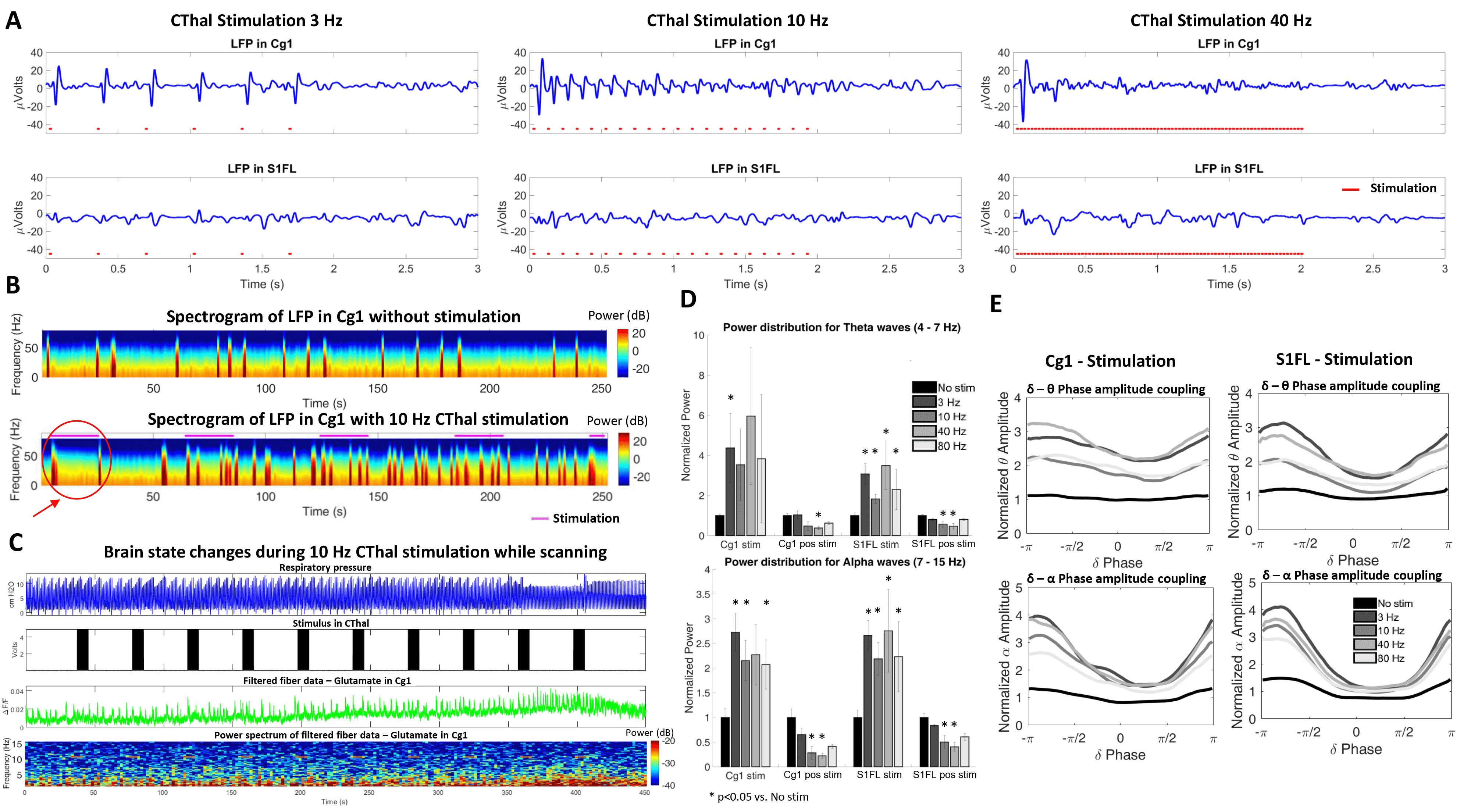
CThal stimulation at the different frequencies triggered a strong positive BOLD response in CThal and in Cg1. However, the different stimulation frequencies triggered varying responses throughout the cortex. 3 Hz stimulation shows the strongest positive BOLD activation in cortical areas, followed by 40 Hz stimulation. This positive BOLD correlation is considerably less with 10 Hz stimulation (Figure 1). This observation suggests that 10 Hz opto-stimulation-driven brain state changes may cause less robust BOLD signal changes in anesthetized rats.In addition, CThal stimulation in deeply anesthetized animals drives the electrophysiological signals in Cg1 and in other regions of the cortex (S1FL) to a profile similar to a less anesthetized (or more awake) state. As the brain transitions to a less anesthetized state, the frequency of bursting events increases, as well as the higher frequency content in the LFP at Cg1 and S1FL (Figure 2A-B). These changes are also observed during the periods of CThal stimulation despite the anesthesia level being maintained (Figure 3A, B and D). Phase amplitude coupling between the delta-theta waves and the delta-alpha waves also increases during the periods of CThal stimulation (Figure 3E), similarly to when animals transition to a less anesthetized state (Figure 2C).
Sustained CThal stimulation in deeply anesthetized animals is occasionally able to drive long-lasting brain state changes similar to transitions to less anesthetized states (which are barely seen in the absence of stimulation). An example of brain state change induced by 10 Hz CThal stimulation is shown in Figure 3C. This change is characterized by an increased number of bursting events, increased high frequency content in the glutamate signal, and changes in the respiratory pattern.
Discussion
Differences observed in BOLD signal activation in the cortex for different CThal stimulation frequencies may be crucial for understanding the triggers of brain state changes to an awake state and to design more effective treatments for disorders of consciousness. Future studies will be directed towards investigating the reasons behind the absence of strong positive BOLD correlation under the 10 Hz stimulation, which may be due to occasional triggering of negative BOLD in the cortex2. This would be similar to what spontaneously happens in awakening humans3. In addition, the potential of these CThal stimulation strategies to promote subcortical reactivation and acute coma recovery will be further studied in our rodent model4.Conclusions
CThal stimulation is capable of triggering brain state changes and electrophysiologically activating the cingulate cortex and other areas of the cortex similar to a more conscious state. BOLD signal activation in the cortex seems to be dependent on the CThal stimulation frequency. These findings provide useful insights into the understanding of frequency-dependent central thalamus-mediated brain state changes and offer the opportunity to evaluate the potential of these stimulation strategies in promoting subcortical reactivation and acute coma recovery in our rodent model.Acknowledgements
This research was funded by NIH funding (RF1NS113278, RF1NS124778, R01NS122904, R01NS120594, R21NS121642), NSF grant 2123971, and the S10 instrument grants (S10 RR023043 and S10 RR023401) to Martino’s Center.References
1. Thengone DJ, Voss HU, Fridman EA, Schiff ND. Local changes in network structure contribute to late communication recovery after severe brain injury. Sci Transl Med. 2016;8(368):368re5. doi:10.1126/scitranslmed.aaf6113. PubMed PMID: 27928029.
2. Liu J, Lee HJ, Weitz AJ, Fang Z, Lin P, Choy M, Fisher R, Pinskiy V, Tolpygo A, Mitra P, Schiff N, Lee JH.Frequency-selective control of cortical and subcortical networks by central thalamus. Elife. 2015;4:e09215. doi:10.7554/eLife.09215. PubMed PMID: 26652162; PMCID: PMC4721962.
3. Olbrich S, Mulert C, Karch S, Trenner M, Leicht G, Pogarell O, Hegerl U. EEG-vigilance and BOLD effect during simultaneous EEG/fMRI measurement. NeuroImage. 2009;45(2):319-32. doi:10.1016/j.neuroimage.2008.11.014. PubMed PMID: 19110062.
4. Pais-Roldan P, Edlow BL, Jiang Y, Stelzer J, Zou M, Yu X. Multimodal assessment of recovery from coma in a rat model of diffuse brainstem tegmentum injury. NeuroImage. 2019;189:615-30. doi:10.1016/j.neuroimage.2019.01.060. PubMed PMID: 30708105.
Figures