3426
Cortical and subcortical proprioceptive contribution to oculomotor control in humans1School of Psychology and Neuroscience, University of St. Andrews, St. Andrews, Scotland, 2Imaging Centre of Excellence, University of Glasgow, Glasgow, Scotland, 3School of Psychology and Neuroscience, University of Glasgow, Glasgow, Scotland
Synopsis
Keywords: fMRI Acquisition, High-Field MRI, Neuroscience
Motivation: The role of proprioception in controlling the movement of the eyes is currently unknown.
Goal(s): To investigate cortical and subcortical activity in response to a proprioceptive stimulation task.
Approach: 7T fMRI was used to measure BOLD signal in the whole cortex and the brainstem while volunteers performed a previously published proprioceptive task.
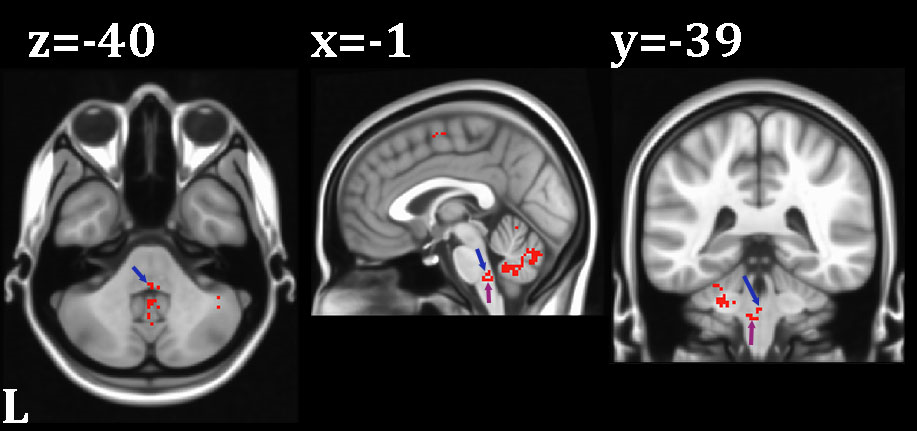
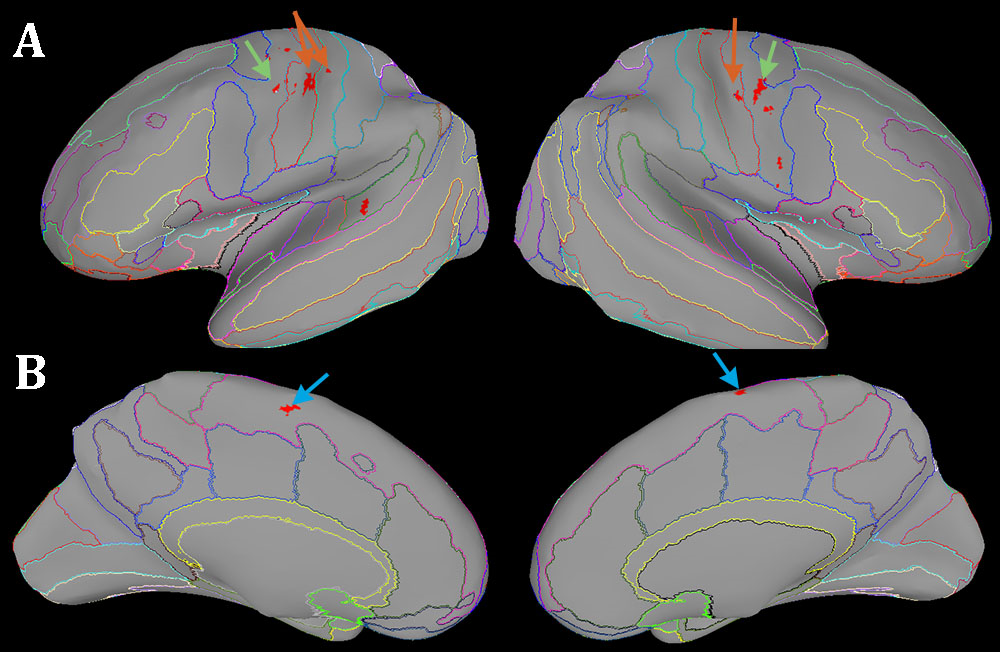
Results: BOLD response was measured in the spinal trigeminal nucleus and the central sulcus/postcentral gyrus, as well as the abducens nuclei, frontal eye fields, supplementary eye fields and the cerebellum.
Impact: A better understanding of the role of proprioception in eye movement control, can open up new avenues for research into the neural mechanisms underlying disorders of eye position and movement like childhood strabismus.
Background
Stretch receptors within the extraocular muscles convey information to the central nervous system about the rotation of the eyes. While precise control of limb position critically relies on proprioceptive feedback, a role for proprioception in controlling eye movements remains uncertain. To investigate whether the oculomotor network in the human brain respond to proprioceptive feedback, we acquired blood oxygen level dependent (BOLD) signal using ultra-high-field functional magnetic resonance imaging (7T fMRI), with a sequence optimized to focus on subcortical activation(1). Previous studies conducted at lower magnetic field strength (3T) identified bilateral activity in the central sulcus (area 3A) and premotor cortex(2). An unexpected finding was that the brainstem’s extraocular motor nuclei that move the left eye responded to proprioceptive stimulation of the right eye’s extraocular muscles(3). We aimed to replicate those findings.Methods
Healthy adult volunteers (N=6) were asked to close their eyes and place their right index finger on the outer corner of their right eyelid. Following an auditory cue, they gently and briefly pushed the eyeball towards the nose, passively stretching the right lateral rectus muscle. Control conditions were designed to isolate motor and tactile task components. There were four conditions. Active: active eye movement; Passive: brief press (< 1 second) at the right corner of the right eye with their right index finger so that to gently move the eyeball. Touch: touch on the eyelid with their index finger, without moving the eyeball and Rest. Trials of each type were grouped in 25s blocks. Neural activity in response to eye proprioception was identified using the conjunction (Active – Rest) AND (Passive – Rest) masked exclusively with (Touch – Rest). The threshold for the conjunction was p<0.05 FDR-corrected for multiple comparisons, whereas for the exclusive mask it was more liberal (p<0.05, uncorrected). This contrast ruled out the confounding effects of finger movement or tactile stimulation on the eyelid. The task and the contrast were as described previously for 3T(3).All imaging was acquired using a 7T MAGNETOM Terra MRI scanner (Siemens, Erlangen, Germany) and single transmit, 32-channel receive radiofrequency head coil (Nova Medical Inc., Wilmington, MA, USA) with local ethical approval. Dielectric pads were used to improve the B1+ homogeneity(4) with additional foam padding used to limit head movement. Functional data were acquired using a multi-band 2D echo-planar interleaved imaging (EPI) sequence(5) with left to right phase-encoding and the following imaging parameters: 144 dynamics, resolution = 2 mm isotropic, 62 slices, field of view (FOV) = 192 x 192 x 124 mm, repetition time (TR) = 2500 ms, echo time (TE) = 17 ms(1), flip angle = 72°, multiband acceleration = 2. A short 2D-EPI scan (5 volumes) was acquired with the opposite phase encoding direction to correct for nonlinear geometric distortions.
After standard preprocessing (slice timing, realignment, distortion correction, normalisation to MNI space and smoothing with FWHM= 2mm), data were analysed using a classic general linear model (3dDeconvolve in AFNI(6, 7)).
Results and Discussion
The stretch of the right lateral rectus muscle was associated with suprathreshold activity not only in the somatosensory but also in the oculomotor network. In the brainstem we found a response to proprioceptive stimulation in the left abducens nucleus and left trigeminal nucleus which send motor commands and receive proprioception, respectively, to/from the extraocular muscles of the left eye. This suggests a proprioceptive interaction between the two eyes. We replicated the cortical activation identified previously(2) and found additional foci in the cerebellum and supplementary eye fields (Figure 1-2).Conclusion
This study replicates and extends previous findings at 3T. It also demonstrates the feasibility of simultaneous whole brain and brainstem 7T fMRI. By seeking to identify the neural connections that support the proprioceptive coupling between the movement of the two eyes, the causes of childhood strabismus, which are often unknown, can be investigated.Acknowledgements
No acknowledgement found.References
1. de Hollander G, Keuken MC, van der Zwaag W, Forstmann BU, Trampel R. Comparing functional MRI protocols for small, iron-rich basal ganglia nuclei such as the subthalamic nucleus at 7 T and 3 T. Hum Brain Mapp. 2017;38(6):3226-48.
2. Balslev D, Albert NB, Miall C. Eye muscle proprioception is represented bilaterally in the sensorimotor cortex. Hum Brain Mapp. 2011;32(4):624-31.
3. Balslev D, Mitchell AG, Faria PJM, Priba L, Macfarlane JA. Proprioceptive contribution to oculomotor control in humans. Hum Brain Mapp. 2022;43(16):5081-90.
4. Teeuwisse WM, Brink WM, Webb AG. Quantitative assessment of the effects of high-permittivity pads in 7 Tesla MRI of the brain. Magn Reson Med. 2012;67(5):1285-93.
5. Moeller S, Yacoub E, Olman CA, Auerbach E, Strupp J, Harel N, et al. Multiband multislice GE-EPI at 7 tesla, with 16-fold acceleration using partial parallel imaging with application to high spatial and temporal whole-brain fMRI. Magn Reson Med. 2010;63(5):1144-53.
6. Cox RW. AFNI: software for analysis and visualization of functional magnetic resonance neuroimages. Comput Biomed Res. 1996;29(3):162-73.
7. Saad ZS, Reynolds RC. Suma. Neuroimage. 2012;62(2):768-73.
8. Duvernoy H. The human brainstem and cerebellum : surface, structure, vascularization and three dimensional sectional anatomy with MRI: Springer-Verlag; 1995.
9. Linzenbold W, Lindig T, Himmelbach M. Functional neuroimaging of the oculomotor brainstem network in humans. Neuroimage. 2011;57(3):1116-23.
10. Schulte LH, Sprenger C, May A. Physiological brainstem mechanisms of trigeminal nociception: An fMRI study at 3T. Neuroimage. 2016;124(Pt A):518-25.
11. Stankewitz A, Voit HL, Bingel U, Peschke C, May A. A new trigemino-nociceptive stimulation model for event-related fMRI. Cephalalgia. 2010;30(4):475-85.
12. Desikan RS, Segonne F, Fischl B, Quinn BT, Dickerson BC, Blacker D, et al. An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. Neuroimage. 2006;31(3):968-80.
13. Glasser MF, Coalson TS, Robinson EC, Hacker CD, Harwell J, Yacoub E, et al. A multi-modal parcellation of human cerebral cortex. Nature. 2016;536(7615):171-8.
Figures