3409
Using saturation bands to null signal from inflowing blood in single-slice fMRI: Toward a rapidly sampled black-blood functional contrast1Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital, Boston, MA, United States, 2Radiology, Harvard Medical School, Boston, MA, United States, 3Center for Advanced Imaging, University of Queensland, St Lucia, Australia, 4Harvard-MIT Division of Health Sciences and Technology, Boston, MA, United States
Synopsis
Keywords: fMRI Acquisition, New Signal Preparation Schemes, Acquisition Methods, Blood Vessels, Brain, Contrast Mechanisms, fMRI (task based), fMRI Acquisition, New Signal Preparation Schemes, Novel Contrast Mechanisms, Pulse Sequence Design, RF Pulse Design & Fields, Vascular, Velocity & Flow, inflow, black-blood contrast, blood nulling, functional contrast, blood flow, blood volume, arteries, veins, flow related signal enhancement
Motivation: Magnetization-preparation schemes using inversion recovery measure aspects of hemodynamics like perfusion and blood volume, but their temporal resolution is limited by physiology, i.e., long blood arrival/transit times.
Goal(s): To study brain hemodynamics at high temporal resolutions with novel inflow saturation.
Approach: We applied saturation bands on both sides of a rapidly-sampled single imaging slice to suppress inflow-related signal and produce black-blood functional contrast sensitive to macrovascular blood volume.
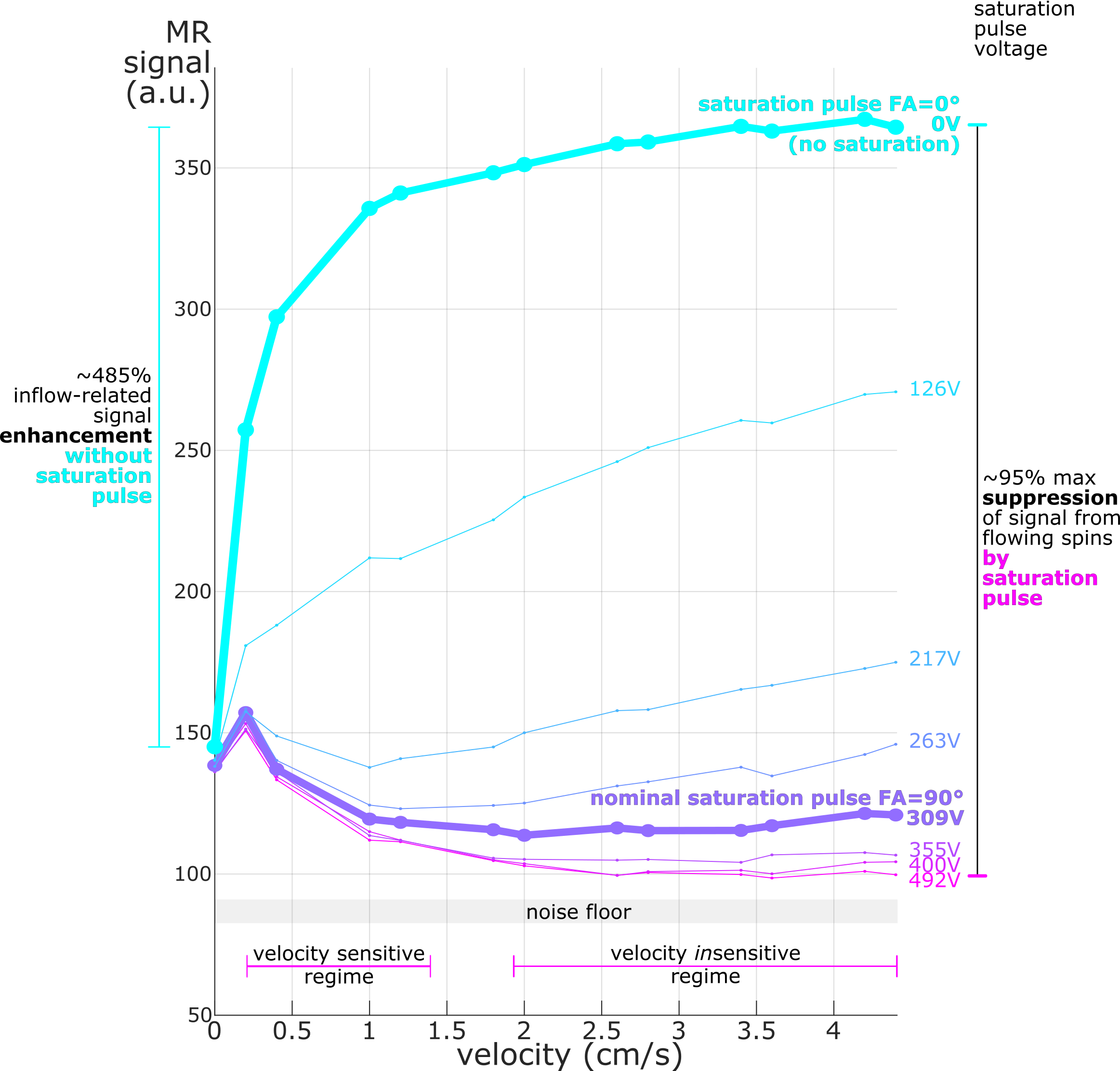
Results: Phantom experiments show successful suppression of inflow signal with flow-velocity-dependence at low- and high-velocity regimes but independence at mid-regime. Human brain vessels exhibited partial signal suppression, with more functional suppression seen in larger vessels.
Impact: Fast blood nulling may be achievable using local saturation bands contiguous to an imaging slice rather than a global inversion slab, enabling functional contrasts sensitive to macrovascular blood flow-velocity and volume suitable for studying fast hemodynamics in specific vascular compartments.
Introduction
Hemodynamic responses to neuronal activity are remarkably precise in space and time, however the interpretation of human fMRI is limited by our incomplete understanding of the link between neuronal and vascular dynamics. Neuronal activity triggers rapid, active vascular responses in individual arterioles then, according to current models, a perhaps slower, passive response occurs in downstream venules. A more complete picture of hemodynamics in humans will require compartment-specific measures of arterial and venous responses at high spatial and temporal resolutions. Here we propose to isolate these compartmental responses using a novel blood-nulling approach.Existing non-BOLD measures of blood flow and volume achieve hemodynamic specificity using blood-pool contrast agents or blood magnetization preparation or “labeling” techniques typically using inversion recovery1–3. However, techniques like ASL or VASO have intrinsically limited temporal resolution: both label blood globally (e.g., in feeding arteries) and then wait for (a) labeled blood to reach the microvasculature and (b) recover to a null longitudinal magnetization. These limits can be circumvented by labeling through saturation bands adjacent to a single imaging slice, thus immediately nulling inflowing blood and providing macro- and perhaps meso-scale vessel specificity while sacrificing capillary weighting and spatial coverage. Although labeling of “proximal” spins was proposed previously for inversion recovery4,5, nulling through saturation enables faster sampling and is compatible with “single-vessel fMRI”6–9 to investigate vessel-compartment-specific dynamics.
Methods
Phantom and human MRI experiments were conducted at 7T (Siemens Magnetom Terra), using an inhouse-built 64-channel receive-coil head array10. To achieve saturation bands with sharp profiles, we added optimized parallel saturation bands to a multi-echo FLASH sequence. Briefly, we designed gradient-spoiled slab-selective RF pulses for saturation (Shinnar-LeRoux, time-bandwidth product=21.1, duration=9.2ms, nominal FA=90°) using Stanford RF toolbox11. Saturation pulses were played Δt=9ms (center-to-center) before the excitation used for imaging. RF spoiling was used for both saturation and imaging pulses. To reduce SAR, we lengthened saturation TR (TRsat) by regularly omitting the pulse while keeping imaging repetition time (TRexc) constant.We evaluated saturation performance on a flow phantom that produces accurate water velocities (Figure1A). The saturation pulse (TRsat=190ms) targeted a 10-mm slab (Figure1C) in front of a single 1.2-mm-width imaging slice (Figure1B; FA=20°, TRexc=19ms, TE=3.1ms, FoV=132mm, 1x1x1.2mm3 voxels, GRAPPA=4, temporal resolution=0.627s) with a nominal 2-mm gap. Saturation efficiency was manipulated by altering RF pulse voltage. Signal was averaged within the flow tubes (Figure 1B) after discarding transients between velocity steps.
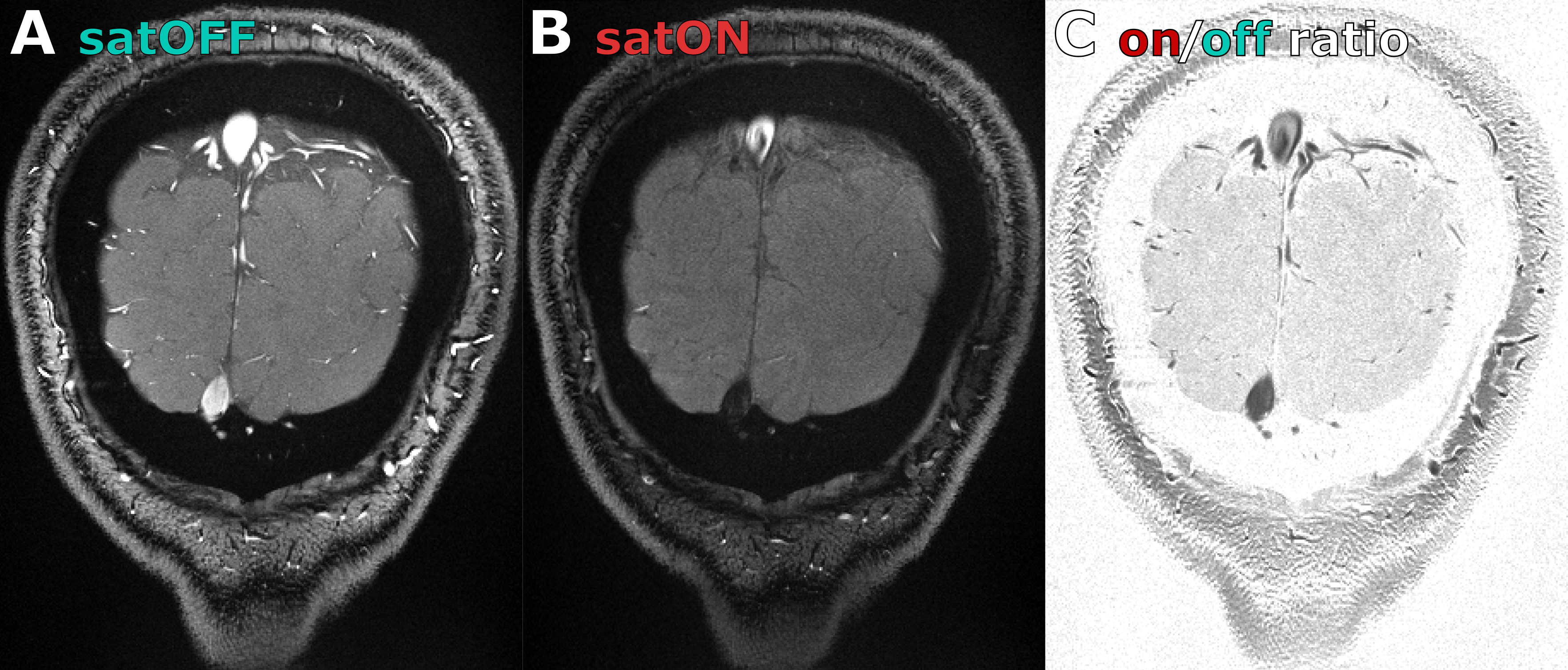
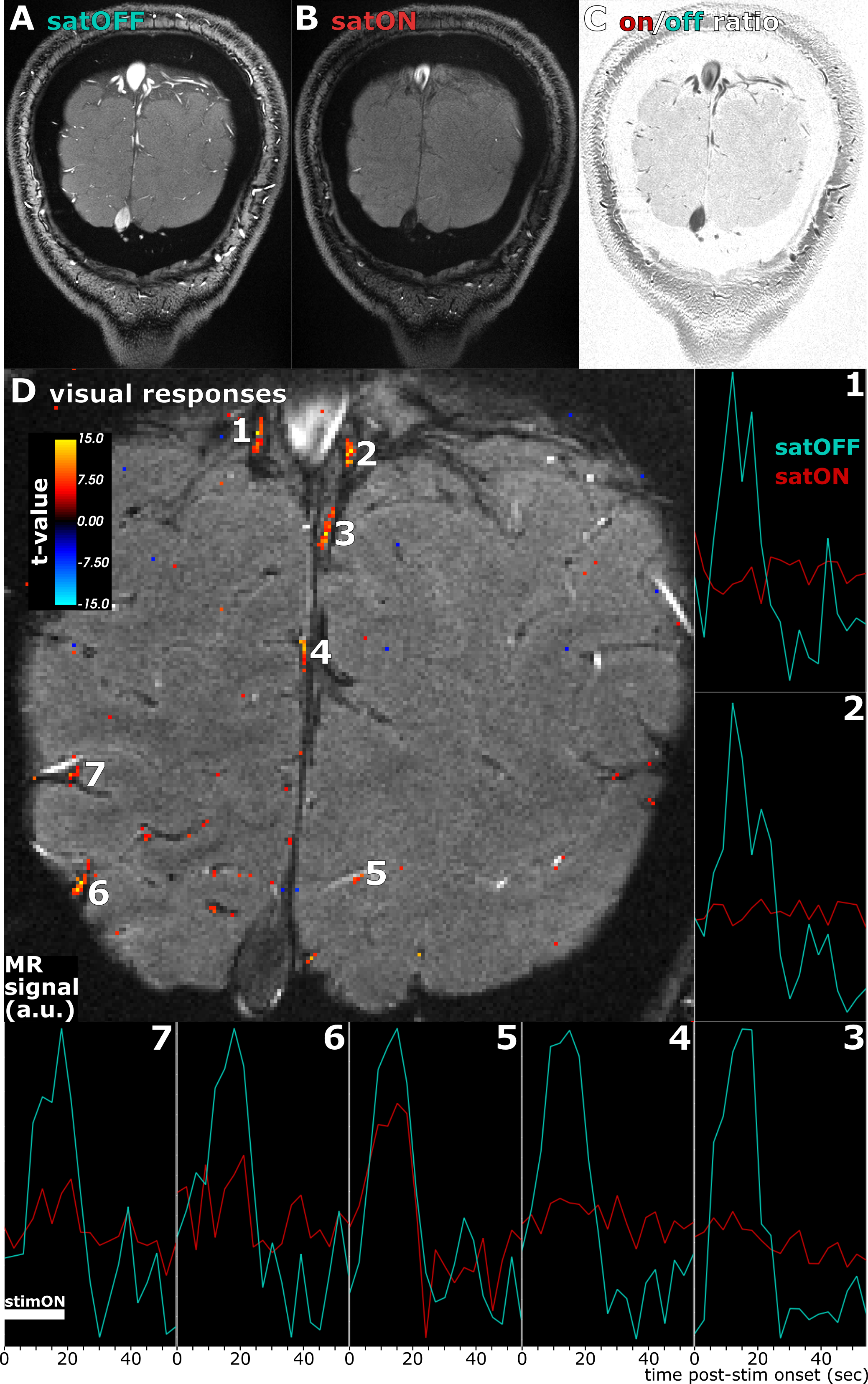
A healthy volunteer (29 y.o. male) was scanned after providing written informed consent, following all policies of our institution’s Human Subjects Research Committee. The same sequence was used with higher spatial resolution and a bipolar multi-gradient-echo readout (FA=26°, TR=30ms, TE=3.6, 6.5, 9.3, 12.1ms, FoV=160mm, 0.4x0.4x1.2mm3 voxels, GRAPPA=4, temporal resolution=3.0s). Saturation was made symmetric by adding an extra saturation pulse targeting the other side of the imaging slice (TRsat=180ms for each side). Saturation efficiency was visualized through saturation ON/OFF signal ratio, averaged across all functional runs.
We measured responses to flashing (8Hz) checkerboard visual stimulation (18s-ON/39s-OFF) during a fixation/attentional task in a coronal slice perpendicular to the calcarine sulcus. Totals of 8 and 16 trials were collected in 2 and 4 runs with saturation OFF and ON, respectively. Across-run motion was compensated prospectively12 and in-plane within-run motion corrected retrospectively. Trial-triggered average responses were estimated on detrended timeseries and significant stimulus ON vs. OFF activations detected with paired t-tests across trials.
Results
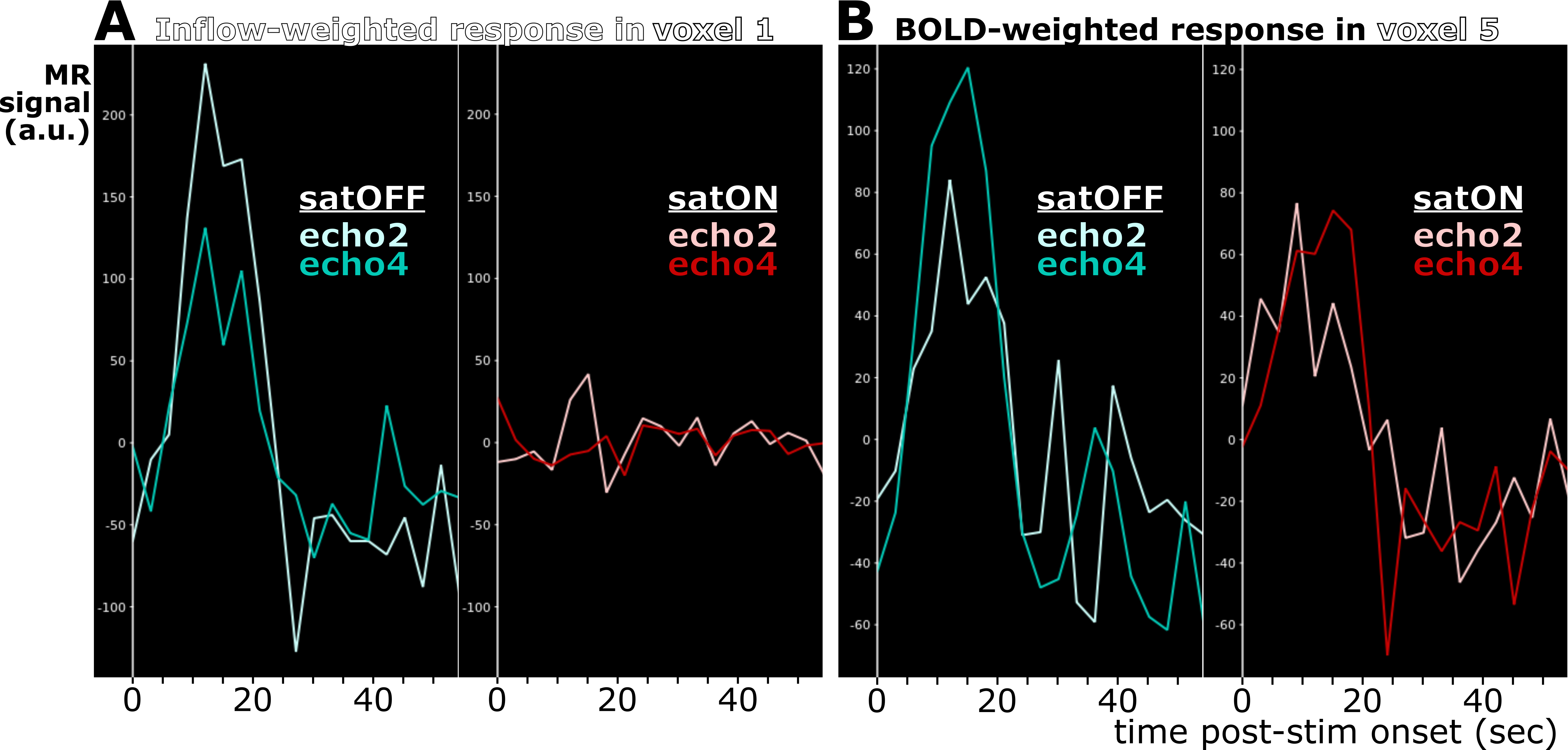
We confirmed strong and specific suppression of signal from spins flowing through the saturation slice and into the imaging slice both in the phantom (Figure1D and Figure2) and in vivo (Figure3). Saturation virtually eliminated functional responses in the large vessels 1 and 2, but partially suppressed responses in and around smaller vessels (Figure4). Responses in vessels 1 and 2 showed evidence of inflow weighting, while the largely preserved response around Vessel 5 was BOLD weighted14 (Figure5).Discussion
Our novel fMRI method for saturating inflow reduced intravascular signal and partially suppressed fMRI responses, but did not fully null vessels. While imperfect RF transmit calibration may be involved, B1+ errors did not appear related to saturation performance (data not shown). Weaker suppression in smaller vessels suggests that slower blood significantly recovers from saturation, concordant with the velocity-sensitive regime (Figure2) observed at low-velocities in the phantom. Together with the velocity-insensitive regime observed at higher velocities, these results raise the prospect of a velocity- and volume-weighted functional contrast from mesoscale vessels that becomes more volume-specific at macroscale. Further optimization of the saturation and imaging pulse profiles, flip angles and duty cycles may allow reaching the full blood-nulling required for fast, black-blood contrast fMRI.Acknowledgements
We thank Estee Perelgut, Sarah Richter and Kyle Droppa for their help with subject recruitment and MRI scanning support, Azma Mareyam for 7T hardware support. Thanks to Dr. Laura Lewis, Baarbod Ashenagar and Amelia Strom for lending equipment for the flow phantom and for help with phantom setup and operation. Thanks to Drs. Paul Wighton and Andre van der Kouwe for sharing their prospective slice-prescription update method.This work was supported in part by the NIH NIBIB (grants P41-EB030006, R01-EB019437 and R01-EB032746), NCCIH (grant R01-AT011429), by the BRAIN Initiative (NIH NIMH grant R01-MH111419 and NIH NINDS grants U19-NS123717 and U19-NS128613), and by the MGH/HST Athinoula A. Martinos Center for Biomedical Imaging; and was made possible by the resources provided by NIH Shared Instrumentation Grant S10-OD023637.
References
1. Lu H, Golay X, Pekar JJ, Van Zijl PCM. Functional magnetic resonance imaging based on changes in vascular space occupancy. Magn Reson Med. 2003;50(2):263-274. doi:10.1002/mrm.10519
2. Huber L, Ivanov D, Handwerker DA, et al. Techniques for blood volume fMRI with VASO: From low-resolution mapping towards sub-millimeter layer-dependent applications. NeuroImage. 2018;164:131-143. doi:10.1016/j.neuroimage.2016.11.039
3. Wong EC, Buxton RB, Frank LR. Implementation of quantitative perfusion imaging techniques for functional brain mapping using pulsed arterial spin labeling. NMR Biomed. 1997;10(4-5):237-249. doi:10.1002/(SICI)1099-1492(199706/08)10:4/5<237::AID-NBM475>3.0.CO;2-X
4. Sutton BP, Ouyang C, Ching BL, Ciobanu L. Functional imaging with FENSI: Flow-enhanced signal intensity. Magn Reson Med. 2007;58(2):396-401. doi:10.1002/mrm.21325
5. Shao X, Guo F, Shou Q, et al. Laminar perfusion imaging with zoomed arterial spin labeling at 7 Tesla. NeuroImage. 2021;245:118724. doi:10.1016/j.neuroimage.2021.118724
6. Chen X, Jiang Y, Choi S, et al. Assessment of single-vessel cerebral blood velocity by phase contrast fMRI. Das A, ed. PLoS Biol. 2021;19(9):e3000923. doi:10.1371/journal.pbio.3000923
7. He Y, Wang M, Chen X, et al. Ultra-Slow Single-Vessel BOLD and CBV-Based fMRI Spatiotemporal Dynamics and Their Correlation with Neuronal Intracellular Calcium Signals. Neuron. 2018;97(4):925-939.e5. doi:10.1016/j.neuron.2018.01.025
8. Varadarajan D, Wighton P, Chen J, et al. Measuring individual vein and artery BOLD responses to visual stimuli in humans with multi-echo single-vessel functional MRI at 7T. In: International Proceedings of the 32nd Annual Meeting of ISMRM. ; 2023.
9. Yu X, Nieman BJ, Sudarov A, et al. Morphological and functional midbrain phenotypes in Fibroblast Growth Factor 17 mutant mice detected by Mn-enhanced MRI. NeuroImage. 2011;56(3):1251-1258. doi:10.1016/j.neuroimage.2011.02.068
10. Mareyam A, Kirsch JE, Chang Y, Madan G, Wald LL. A 64-Channel 7T array coil for accelerated brain MRI. In: International Proceedings of the 28nd Annual Meeting of ISMRM. ; 2020.
11. Pauly J, Le Roux P, Nishimura D, Macovski A. Parameter relations for the Shinnar-Le Roux selective excitation pulse design algorithm. IEEE Trans Med Imaging. 1991;10(1):53-65. doi:10.1109/42.75611
12. Van Der Kouwe AJW, Benner T, Fischl B, et al. On-line automatic slice positioning for brain MR imaging. NeuroImage. 2005;27(1):222-230. doi:10.1016/j.neuroimage.2005.03.035
13. Reuter M, Rosas HD, Fischl B. Highly accurate inverse consistent registration: A robust approach. NeuroImage. 2010;53(4):1181-1196. doi:10.1016/j.neuroimage.2010.07.020
14. Havlicek M, Ivanov D, Poser BA, Uludag K. Echo-time dependence of the BOLD response transients – A window into brain functional physiology. NeuroImage. 2017;159:355-370. doi:10.1016/j.neuroimage.2017.07.034Figures