3326
Multidimensional analysis of longitudinal mechanical stimulus-induced fMRI in Mice of both sexes1Clinic of Radiology, University of Münster, Münster, Germany, 2Department of Anesthesiology Intensive Care, Pain Medicine of the University Hospital Münster, Münster, Germany
Synopsis
Keywords: Task/Intervention Based fMRI, fMRI (task based), functional connectivity, acute post-surgical pain, sex difference, mechanosensation, dimensionality reduction, principal component analysis
Motivation: Acute postsurgical pain remains difficult to treat and may result in complications including chronic pain and increased use of opioids if insufficiently controlled.
Goal(s): A better understanding of the mechanisms inducing and maintaining these pain states is critical in both sexes.
Approach: Longitudinal task fMRI was performed with two different mechanical stimulations in male and female mice following acute incision pain (INC) compared to sham-treated animals.
Results: Our mixed-method analysis approach identifies INC-regulated brain regions and highlights divergent mechanical pain processing in mice of both sexes.
Impact: Our mixed-method analysis approach integrating the general linear model analysis combined with principal component analysis and analysis of functional connectivity provides deep insights into CNS plasticity during incision-induced pain and provides detailed information on relevant brain regions and interaction patterns.
Introduction
Acute pain after surgery is still challenging to treat and inadequate control poses a potential risk factor for complications including chronic postsurgical pain and increased use of opioids. Another risk factor for severe pain and chronification after surgery is sex. Therefore, a comprehensive understanding of the sex-specific, cerebral processing of evoked pain after surgery is necessary. Here, we performed task fMRI using two different mechanical stimulation modalities, von Frey (vF) and pinprick (pp), in male and female mice. Measurements were performed in a model of acute post-operative pain1 (incision, INC) and in sham-treated animals (SHAM).Methods
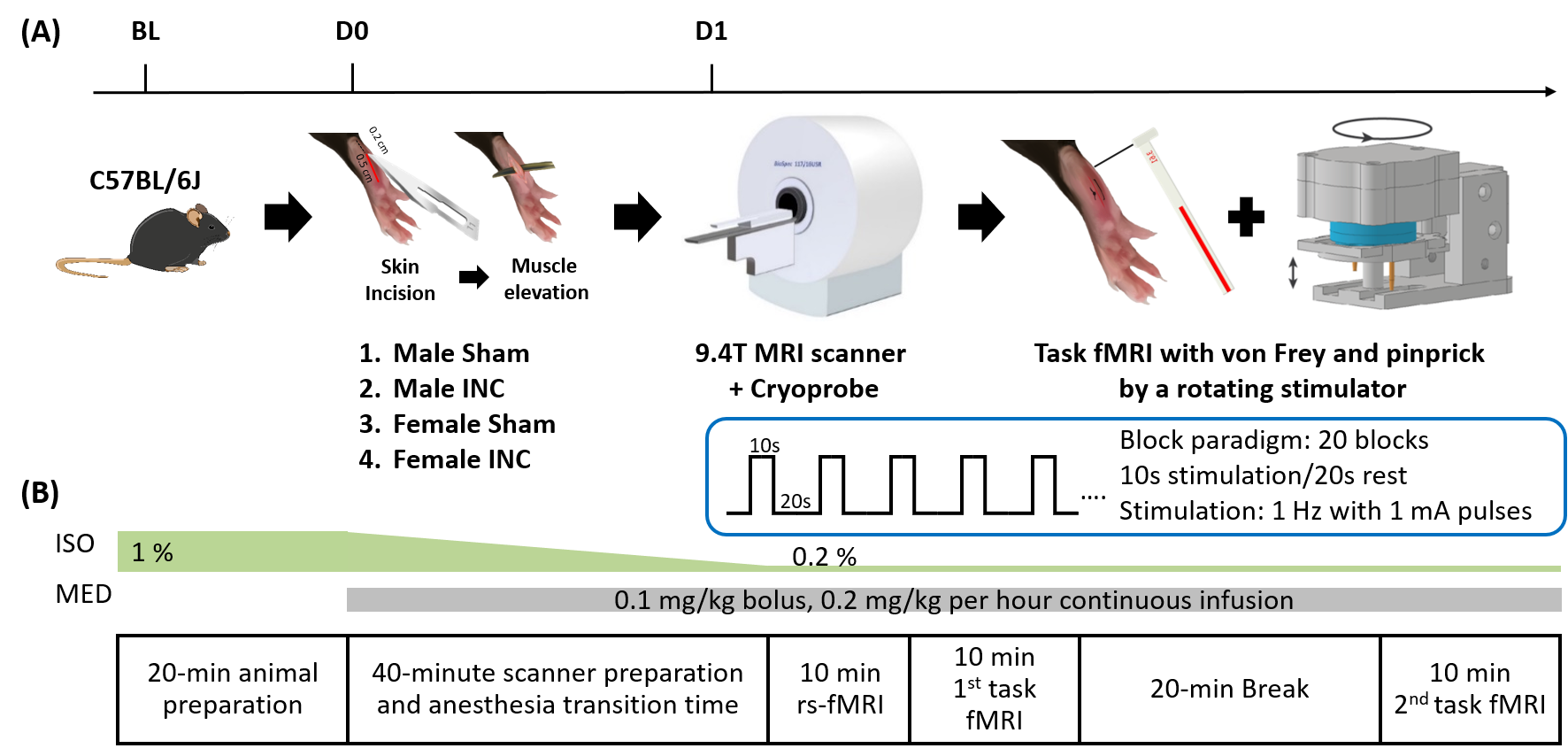
8 to 10-week-old C57BL/6J male and female mice were scanned using a Cryoprobe in a 9.4T MRI scanner as baseline (BL) (Fig. 1). The experiment including two task and one resting-state fMRI measurements was performed using a single-shot gradient EPI sequence (TR/TE 1000/18ms, 18 slices, thickness 0.5 mm, matrix 76x66, resolution 200x200μm2). Mice were anesthetized with mixed isoflurane (0.2%) and medetomidine2. After calibration (20g), noxious (pinprick, pp) and innocuous (von Frey filaments, vF) stimulations were applied using a MR-compatible rotating stimulator on the right hind paw3. Stimulation design was a block paradigm with 20 repetitions of 10s ON / 20s OFF and a stimulation frequency of 1Hz (Fig.1). One day after INC or SHAM procedure (D1) on the right hind paw1, the fMRI experiment was performed again. After preprocessing and general linear model (GLM) analysis with mouse HRF3 using SPM12 and group registration using MagnAn, the BOLD map of individual fMRI data was extracted as a binary file. For determination of functional connectivity (FC)2, the multi-seed region approach (MSRA) was used to acquire averaged correlation matrices followed by calculation of the variance for each functional connection. For statistical evaluation of the short-term modulation of connectivity between different conditions, the network-based statistics (NBS) which is implemented in MagnAn was used with 1000 permutations and pFWE = 0.05.Results
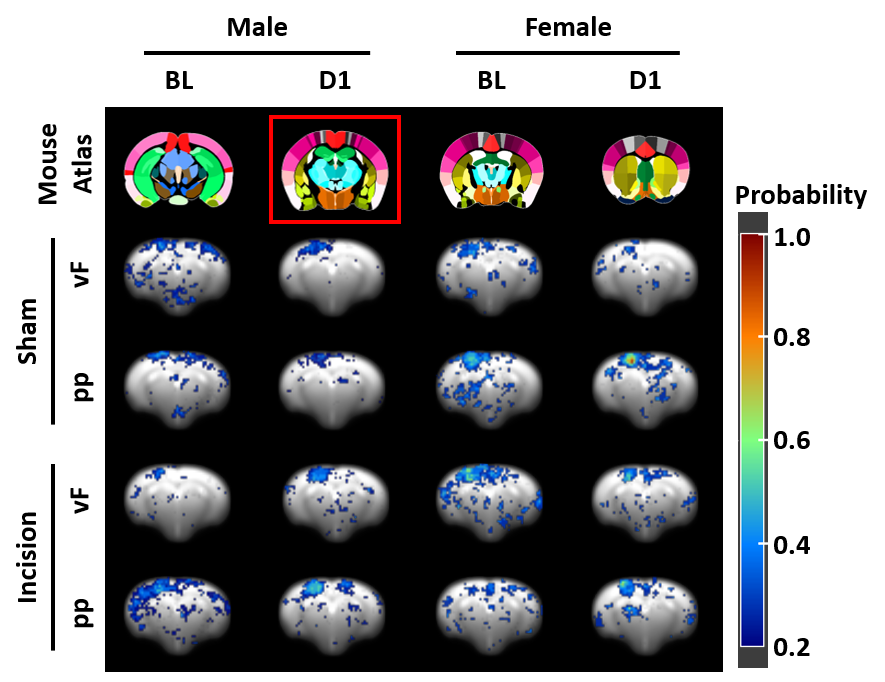
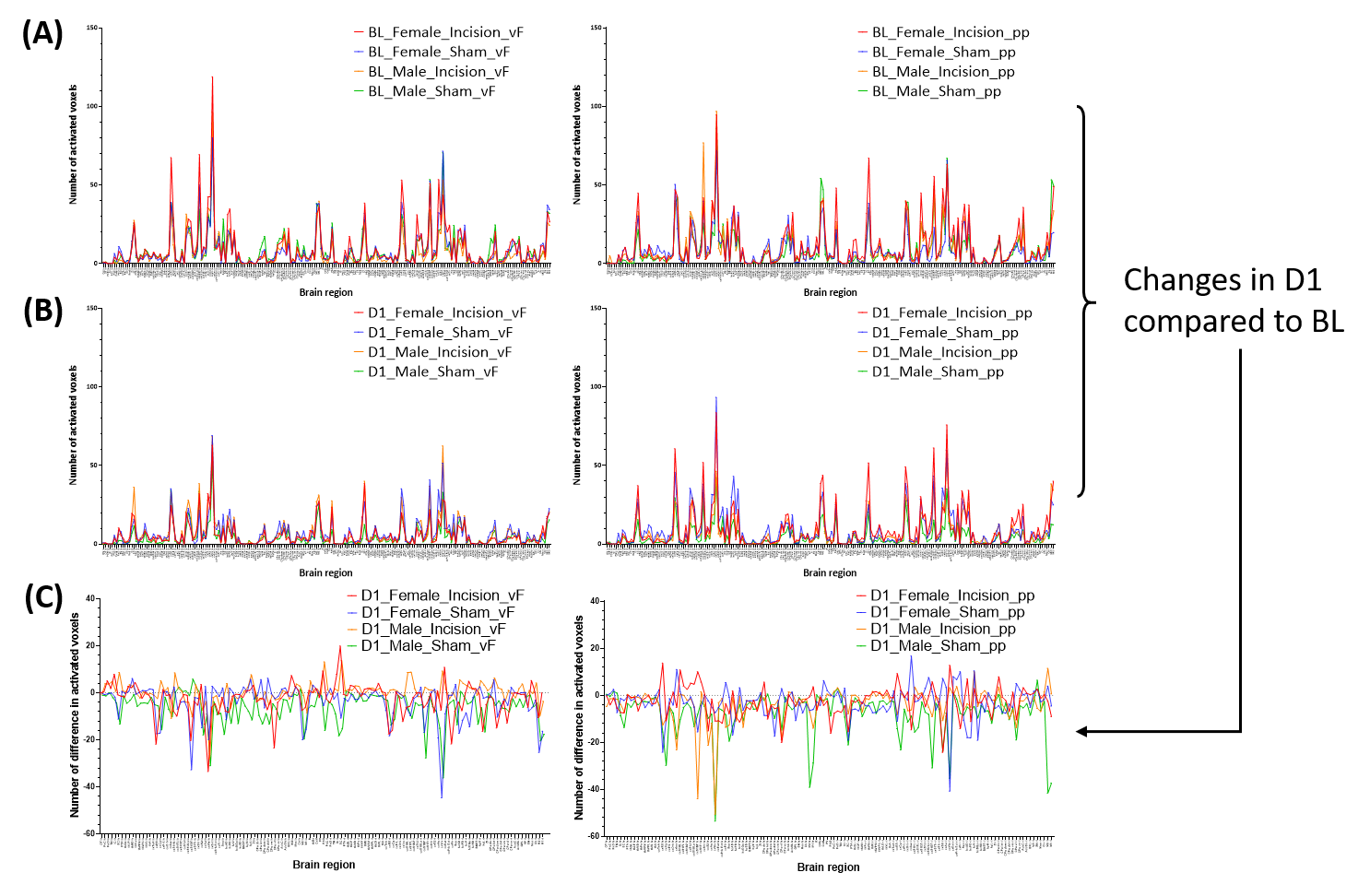
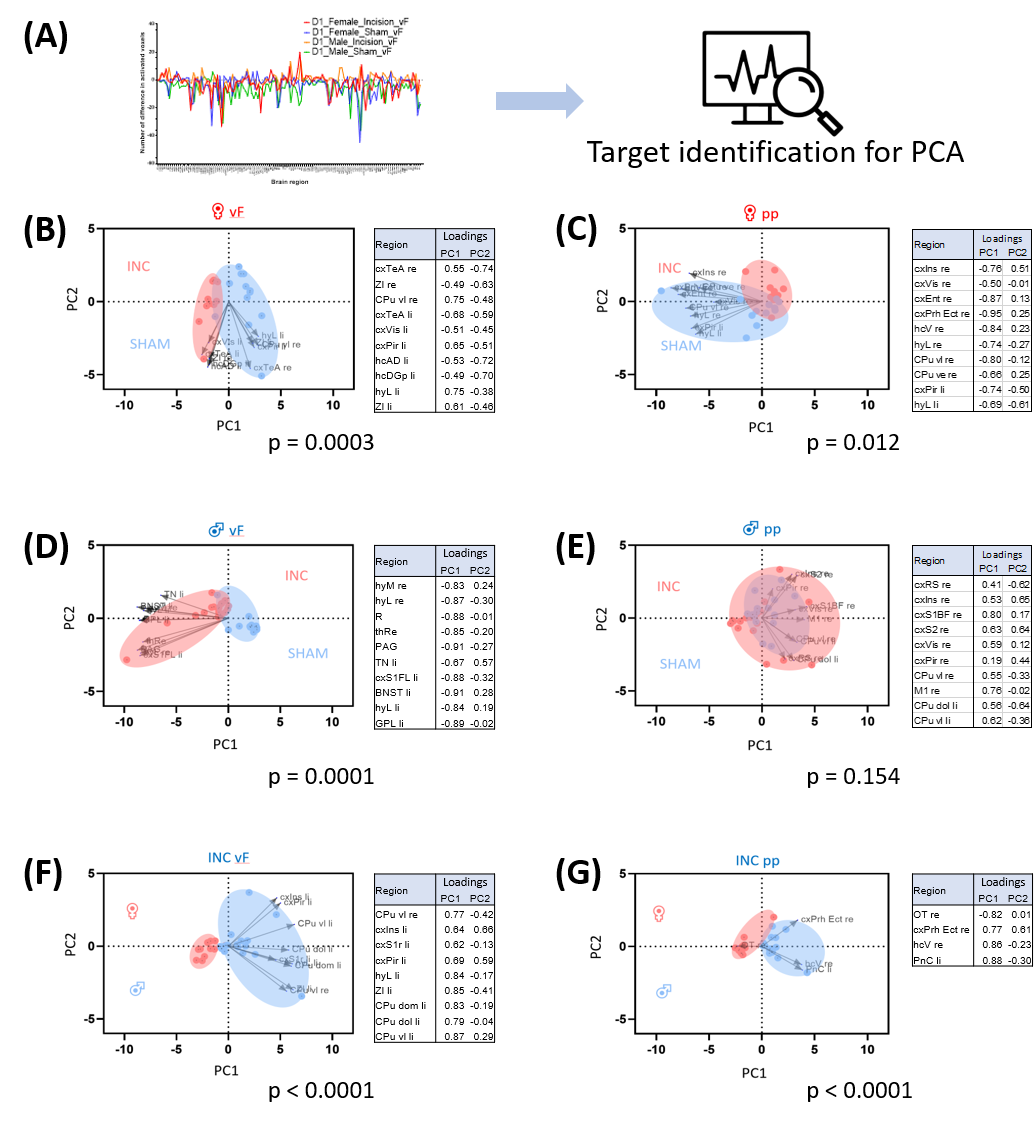
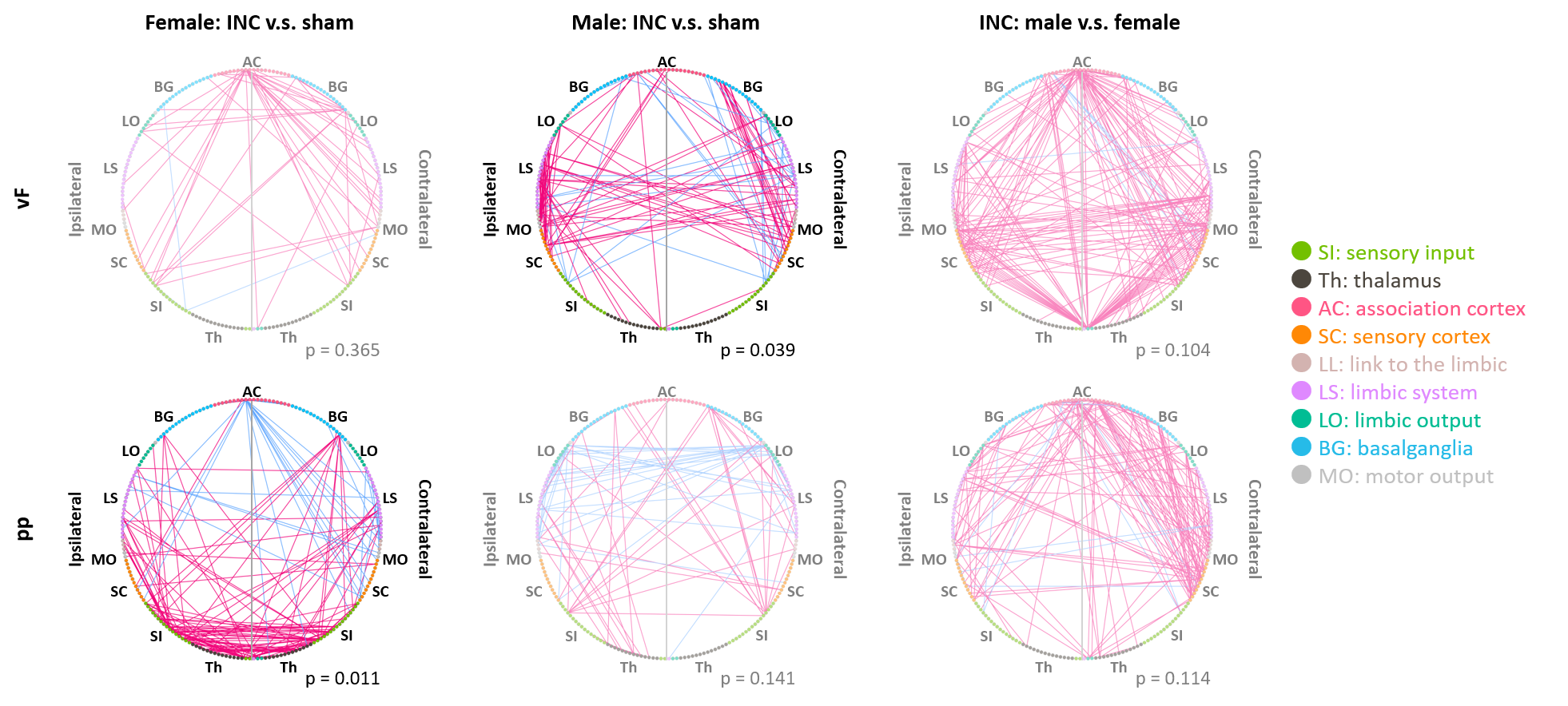
In response to mechanical stimulation with pp and vF, we detected reliable BOLD activation in the S1 hind limb (S1HL) and motor (M1) cortex across all groups (Fig. 2). Consistent with our previous report3, we observed significantly higher numbers of activated voxels in female than in male mice (3-way ANOVA, p<0.05, Fig. 3, top). Following surgery, we found an overall effect of pain model (p<0.001) and sex, with both stimulations separately (p<0.001, Fig. 3 middle). To identify brain regions that showed changes in activation following INC or SHAM procedure (i.e. at D1), we subtracted the mean of each group at BL from the D1 dataset. After excluding regions with differences in less than 4 voxels, 128 brain regions were identified (Fig. 3, bottom). We performed dimensionality reduction using principle component analysis (PCA), starting with regions that showed a mean difference larger 1 standard deviation between any two groups (Fig. 4). With the exception of the male pp group, we found significant differences between INC and SHAM and between male or female groups, based on only 4-10 brain regions. Among these, the most pronounced differences resulting from INC were observed in subdivisions of the hypothalamus (Ht), caudate putamen (CPu) and the rhinal cortex. On the other hand, Ht, CPu, insular and S1 discriminated best between sexes, for animals that had received INC. Next, we analyzed the FC to identify interaction patterns among activated brain regions, and between activated and non-activated regions (Fig. 5). Using the NBS, we found significantly altered whole brain networks between INC and SHAM groups in females following pp, and in males following vF. In females, INC resulted in reorganization of the thalamus, hippocampus and regions integrating sensory inputs. In contrast, INC in males resulted in network modulation of the CPu, amygdala and sensorimotor regions.Conclusion
The combination of GLM and PCA analyses provides complementary results and deep insights into CNS plasticity during incision-induced pain. Following mechanical stimulation, the hind paw incision modulated the descending pain modulatory system, stress response and affective-motivational state in both males and females. While whole-body sensory integration and emotional/sensory processing was increased in males compared to females at D1. Together, these data highlight dynamic neurophysiological adaptations specific to mechanical hypersensitivity during acute post-surgical pain and shed light on the divergent processing of mechanical pain in males and females. Future analyses will further delineate the role of target regions in the brain network.Acknowledgements
No acknowledgement found.References
1. Segelcke, D. et al. Tmem160 contributes to the establishment of discrete nerve injury-induced pain behaviors in male mice. Cell Rep. 37, (2021).
2. Pradier, B. et al. Combined resting state-fMRI and calcium recordings show stable brain states for task-induced fMRI in mice under combined ISO/MED anesthesia. Neuroimage 245, 118626 (2021).
3. Chen, H. et al. Generation of a whole-brain hemodynamic response function and sex-specific differences in cerebral processing of mechano-sensation in mice detected by BOLD fMRI. 1–14 (2023) doi:10.3389/fnins.2023.1187328.
Figures