3325
Astrocytic calcium signal bidirectionally regulated BOLD-fMRI signals in awake mice1Center for Excellence in Brain Science and Intelligence Technology (Institute of Neuroscience), CAS, Shanghai, China
Synopsis
Keywords: Task/Intervention Based fMRI, fMRI, neurovascular coupling
Motivation: Previous study found brain-state dependent astrocytic calcium signals are related to positive and negative BOLD-fMRI signals. However, causal evidences regarding the contribution of astrocytes to BOLD signals is lacking.
Goal(s): We aimed to provide causal evidences for the bidirectional contribution of astrocytic calcium activity on BOLD-fMRI signals.
Approach: We applied simultaneous calcium imaging and awake mouse fMRI to detect distinct astrocytic signals (task-aligned vs. spontaneous) coupled to BOLD signals in three conditions, including no manipulation, activation and inhibition of S1BF astrocytes.
Results: We found whisker stimulation-evoked astrocyte activity mainly contributes to positive BOLD signals, while state-specific astrocyte activity contributes to negative BOLD signals.
Impact: We provide casual evidences that astrocytes bidirectionally regulated BOLD signals. Moreover, our results challenge the prevailing interpretation of the BOLD signal solely as an indicator of neuronal activity by highlighting the impact of arousal-specific involvement of astrocytes in neurovascular coupling.
Introduction
Astrocytes play important roles in the regulation of BOLD signals through the neurovascular coupling1–3. Recently, Wang et al.4 reported that sensory stimulation evoked astrocytic calcium signals were correlated with positive BOLD signals, while state-specific astrocytic calcium signals were correlated with negative BOLD signal in anesthetized rats. However, the causal evidence regarding the contribution of astrocytic calcium activity to the BOLD signal is lacking. In this study, we combined (1) awake mice fMRI with neuronal and astrocytic calcium recording, (2) astrocytic manipulation and (3) video monitoring of behavioral states to elucidate the mechanism of the bidirectional contributions on BOLD responses from stimulus-evoked and state-specific astrocytic calcium signals.Methods
Seven mice were used in each awake mouse fMRI experiment. The viruses used for labeling S1BF neuronal, astrocytic signal or activation/inhibition of the S1BF astrocytes were shown in Main Figures (Fig.1-4), respectively. Optical fiber and head holder implantation were performed one week after virus injection. The spectrally resolved fiber photometry system was shown in Fig. 1, which was used for recording fluorescence signals during the fMRI scanning.All MRI data were collected using multiband EPI sequence with following parameters: TR=750 ms, TE=15 ms, flip angle=46.8°, bandwidth=300 kHz, field of view=15×10.5 mm2, matrix size=100×70, slice thickness=0.48 mm, 1200 volumes per scan, and 38 axial slices per volume. A T2-weighted structural image was acquired for co-registration with following parameters: TR=3400 ms, TE=33 ms, field of view=16×16 mm2, matrix size=256×256, slice thickness=0.5 mm, 30 axial slices, RARE factor=8, and number of averages=2. Each simultaneous fiber photometry and fMRI session included 5 or 8 EPI scans with whisker stimulations and the same number of EPI scans in resting state. In addition, two custom-made MR-compatible video cameras (25 fps) was placed inside the bore to record the mouse spontaneous behaviors.
After basic preprocessing procedures of fMRI data, including rigid-body and nonlinear spatial coregistration and 0.4-mm spatial smoothing, the BOLD signals were further regressed by “6 rp+6 Δrp+10 PCs” nuisance signals to minimize the effects of scanner drift, motion and other non-neural physiological noises adopted from our previous study5, in which “6 rp+6 Δrp” nuisance signals represented 6 head motion parameters and their 1st order first derivatives, and “10 PCs” were the first 10 principal components from the BOLD signals of non-brain tissue, e.g., the muscles.
Results
Fig. 1 showed the timeline of simultaneous fiber photometry and fMRI in awake mice. Gradient air pressure test showed that 16-psi whisker stimulation was sufficient to evoke robust S1BF neural and astrocytic activation (Fig. 1D-E). Combined with spontaneous behavioral monitoring by MR-compatible cameras during fMRI scanning (Fig. 2), we found whisker stimulation accompanied with body movement caused larger astrocytic calcium signal compared with whisker stimulation only (Fig. 2D-E). Such larger astrocytic calcium signal coupled to delayed negative BOLD response (Fig. 2E-F).Thus, we hypothesized that stimulation evoked astrocytic calcium signals contribute to positive BOLD response, while spontaneous state-specific astrocytic calcium signals contribute to negative BOLD response (Fig. 3).
To validate our hypotheses, we firstly examined how the calcium elevation in S1BF astrocytes contributes to BOLD signals by elevating astrocyte calcium with a chemogenetic method, i.e., GfaABC1D-hM3Dq + clozapine (Fig. 4). We found whisker stimulation evoked higher astrocytic and positive BOLD responses in the task-aligned condition. However, in the concurrent (task-aligned and spontaneous) condition, whisker stimulation evoked more apparent increases of ipsilateral neuronal, astrocytic responses by chemogenetic activation of S1BF astrocytes, but no significant increases of ipsilateral positive BOLD signals (Fig. 4C). In the resting state, higher spontaneous astrocytic signal fluctuations induced higher ipsilateral neuronal responses but lower ipsilateral BOLD signal dynamics for both positive and delayed negative ones (Fig. 4D).
Moreover, we performed a loss-of-function experiment by selectively suppressing calcium elevation in S1BF astrocytes using the IP3R2flox mice. Injection of GfaABC1D-Cre into the S1BF of the IP3R2flox mice suppressed astrocyte calcium transients and then we found whisker stimulation evoked lower astrocytic and BOLD responses in both task-aligned only and concurrent conditions (Fig. 5B). In the resting state, inhibition of S1BF astrocytes reduced the amplitude of ipsilateral spontaneous BOLD signal dynamics (Fig. 5C).
Conclusion
In this study, we revealed the coupling between evoked and state-dependent astrocyte calcium signal to positive and negative BOLD signal respectively, and provided causal evidences through the activation and inhibition of S1BF astrocytic signals. These results demonstrated that evoked astrocyte calcium mainly contributes to positive BOLD signal and state-specific astrocyte calcium contributes to negative BOLD signal. Our results made great sense on understanding astrocytes serve distinct roles in directly sensing neuronal activity and feedback to arousal transients through different neurovascular coupling process.Acknowledgements
This work was supported by the National Science and Technology Innovation 2030 Major Program (2021ZD0200100 to ZL), Strategic Priority Research Program of Chinese Academy of Sciences (XDBS01030100 to ZL), Pioneer Hundreds of Talents Program from the Chinese Academy of Sciences (to ZL), Shanghai Municipal Science and Technology Major Project (2018SHZDZX05 to ZL), the National Natural Science Foundation of China (82171899 to ZL), Lingang Laboratory (LG202104-02-06 to ZL), Postdoctoral Innovative Talents Support Program (BX20230383 to CT).References
1. Petzold, G. C. & Murthy, V. N. Role of Astrocytes in Neurovascular Coupling. Neuron 71, 782–797 (2011).
2. Attwell, D. et al. Glial and neuronal control of brain blood flow. Nature 468, 232–243 (2010).
3. Otsu, Y. et al. Calcium dynamics in astrocyte processes during neurovascular coupling. Nat Neurosci 18, 210–218 (2015).
4. Wang, M., He, Y., Sejnowski, T. J. & Yu, X. Brain-state dependent astrocytic Ca 2+ signals are coupled to both positive and negative BOLD-fMRI signals. Proc. Natl. Acad. Sci. U.S.A. 115, (2018).
5. Chen, X. et al. Sensory evoked fMRI paradigms in awake mice. NeuroImage 204, 116242 (2020).
Figures
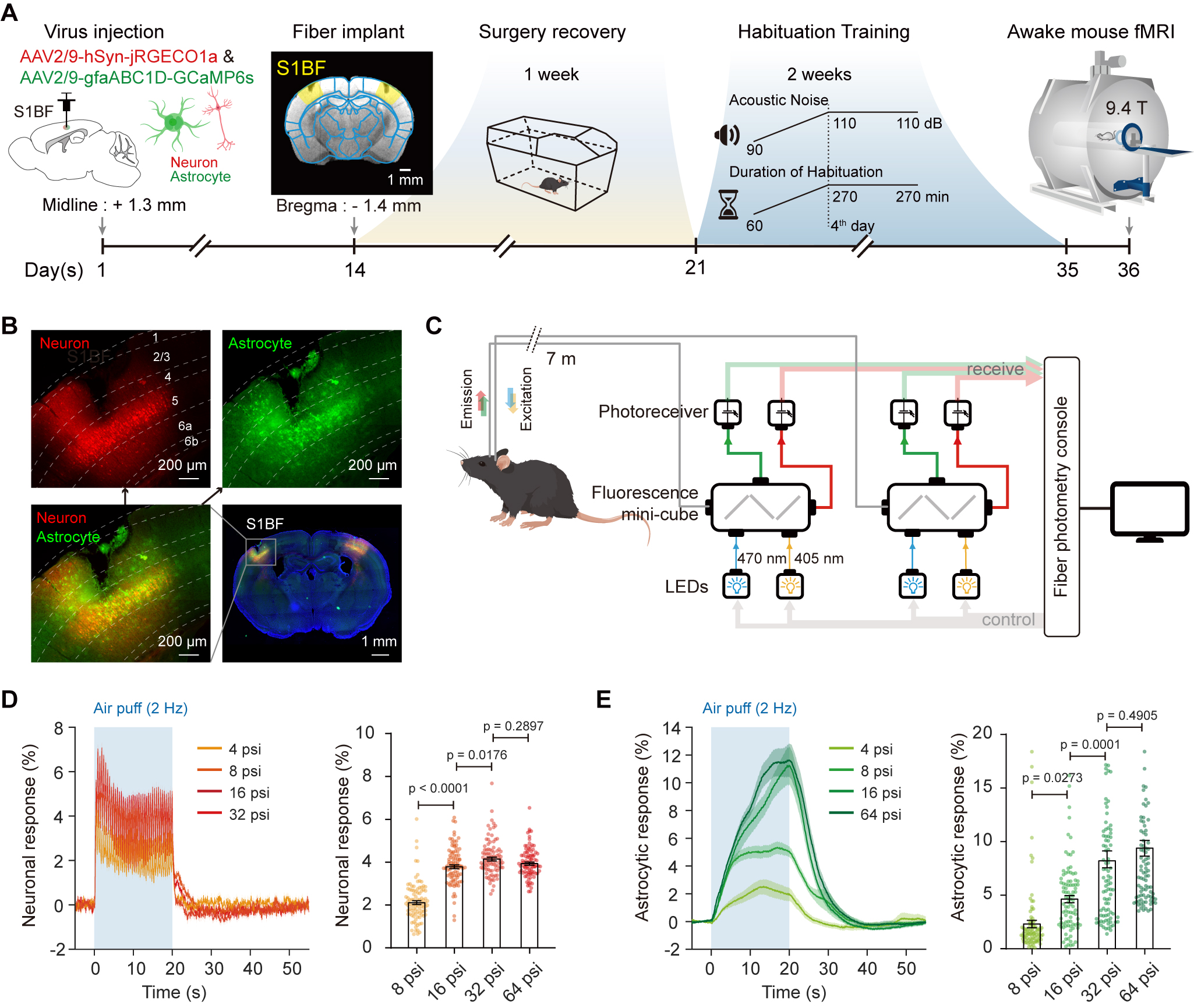
Figure 1. Schematic illustration of the simultaneous calcium fiber photometry and fMRI setup.
(A) Timeline of simultaneous fiber photometry and fMRI in awake mice. (B) Expression of GCaMP6s (for labeling astrocytes) and jRGECO1a (for labeling neurons) in S1BF. (C) Schematic of a spectrally resolved fiber photometry system used for recording fluorescence signals. (D-E) Neuronal (D) and astrocytic (E) response of different air pressures of whisker stimulation.
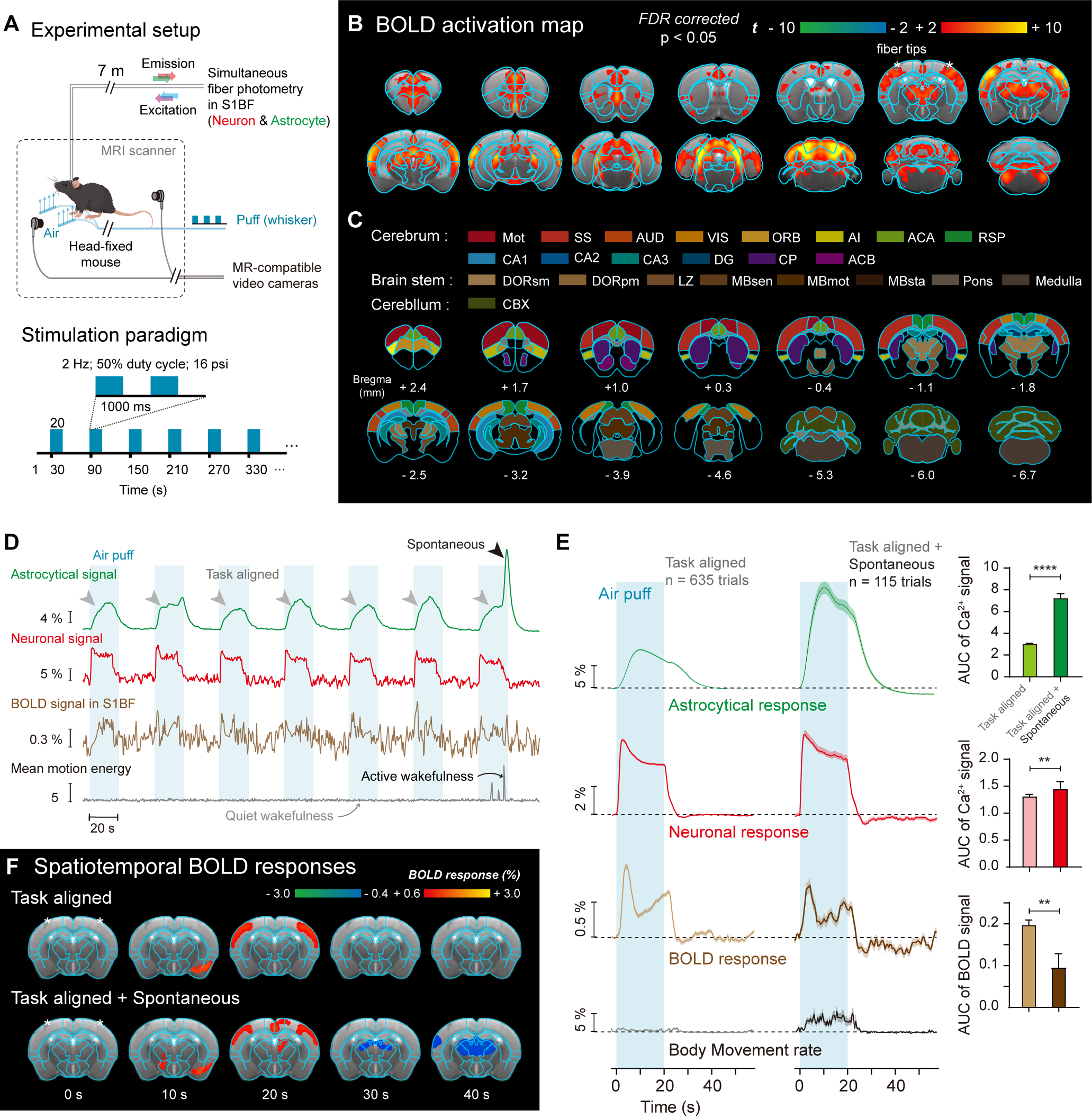
Figure 2. Spontanous state-specific actrocytic calcium signal coupled to negative BOLD response.
(A) Experimental setup of simultaneous calcium fiber photometry and awake mouse fMRI with whisker stimulation. (B) BOLD activation map of whisker stimulation in awake mice. (C) Parcels based on CCFv3 Allen mouse brain atlas. (D-E) Representative and averaged whisker stimulation evoked astrocytic and neuronal calcium, BOLD and mouse behavioral signals in S1BF. (F) Whisker stimulation evoked spatiotemporal maps of BOLD signals.
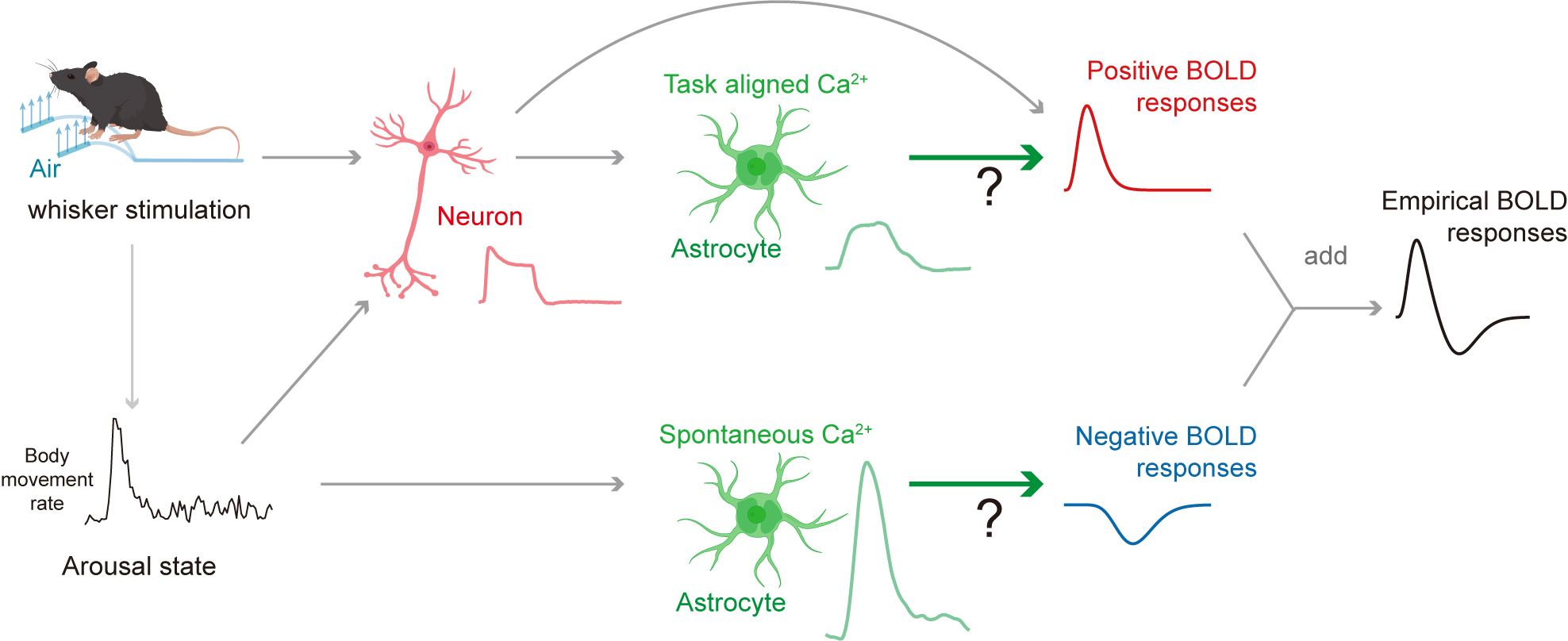
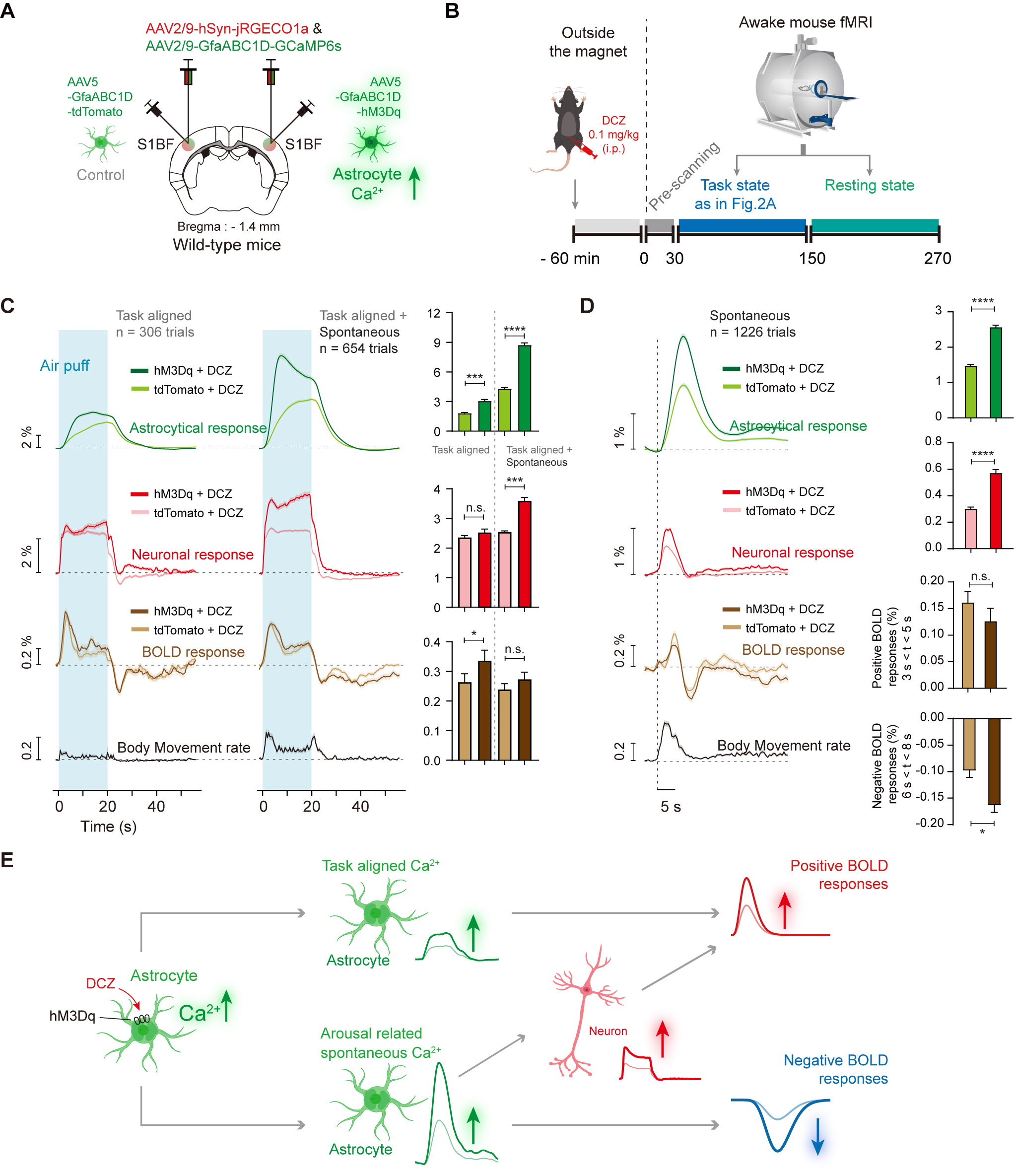
Figure 4. Chemogenetic activation of S1BF astrocytes induced higher BOLD-fMRI signal dynmaics for both positive and negative BOLD-fMRI signals.
(A-B) Schematic diagram depicting the chemogenetic activation of astrocytes and awake mouse fMRI. (C) Chemo-activation of task-aligned astrocytes induced higher positive BOLD responses. (D) Chemo-activation of spontaneous astrocytes signals induced higher negative BOLD responses. (E) Diagram illustrating neuronal, astrocyte and BOLD signal dynamics induced by chemogenetic activation of S1BF astrocytes.
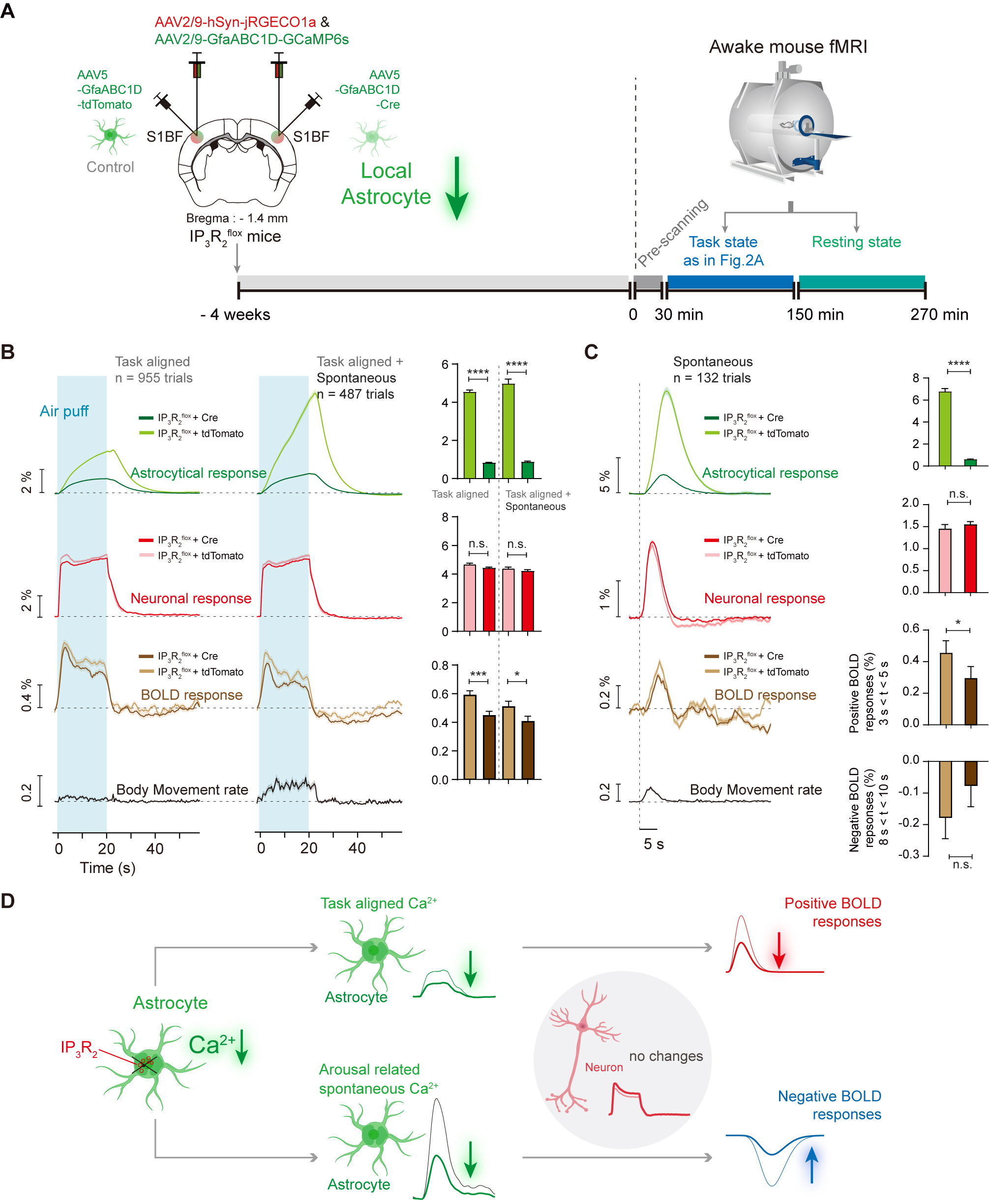
Figure 5. Decreased astrocyte activity in S1BF induced lower BOLD-fMRI signal dynmaics for both positive and negative BOLD-fMRI signals.
(A) Schematic diagram depicting IP3R2 knockout (KO) + GfaABC1D-Cre reduced astrocyte calcium signal for awake mouse fMRI. (B) IP3R2-KO in S1BF astrocytes reduced the task-aligned positive BOLD responses. (C) IP3R2-KO in S1BF astrocytes reduced the amplitude of spontaneous BOLD signal dynamics. (D) Diagram illustrating neuronal, astrocyte and BOLD signal dynamics induced by IP3R2-KO in S1BF astrocytes.