3316
Perturbation of intrinsic oscillatory modes by focal ischemic stroke in rats1Champalimaud Research, Champalimaud Foundation, Lisbon, Portugal, Lisbon, Portugal, 2Life and Health Sciences Research Institute, University of Minho, Braga, Portugal, Braga, Portugal
Synopsis
Keywords: Functional Connectivity, Brain
Motivation: The biophysical mechanisms underlying changes in functional connectivity upon stroke remain unclear.
Goal(s): The present study aims at investigating how disrupting a specific and well-localized cortical area modulates spontaneous long-range functional connectivity.
Approach: A novel approach based on intrinsic oscillatory modes exhibiting stationary wave pattern features was used to provide insight into the organizing principles underpinning spontaneous long-range FC.
Results: The presented intrinsic mode framework revealed global reorganization following local disruption with enhancement of corticostriatal activity in stroke.
Impact: Our findings provide an important insight augur well for better understanding of neural network reorganization.
Introduction
Disruption of functional networks is a hallmark of numerous brain disorders1-3. In stroke, loss of interhemispheric functional connectivity (FC) has been reported at the acute stages4, followed by renormalization at later stages in some cases5. However, the biophysical mechanisms underlying these changes in FC remain to be elucidated. Recently, a repertoire of intrinsic oscillatory modes exhibiting stationary wave pattern features were discovered in healthy animal brains and in humans6 and provided insight into the organizing principles underpinning spontaneous long-range FC7. Here, we investigated how disrupting a specific and well-localized cortical area modulates these intrinsic modes.Methods
All animal experiments were preapproved by the competent national and international authorities and were carried out according to EU Directive 2010/63. Adult Long Evans rats female were used (N = 20) weighing 350+/-100g.Stroke induction: A photothrombotic stroke8 was induced in M1 of the left hemisphere (N = 10) by injecting Rose Bengal dye (Sigma Aldrich, Portugal) (15 mg/ml) delivered intravenously (10 μl/g) followed by 15min irradiation with a cold light source8. Stroked animals (N = 10) were imaged 1w poststroke along with N=10 healthy controls, under medetomidine sedation while monitoring temperature and respiration rate.
MRI: 9.4T BioSpec scanner (Bruker, Germany) with an 86mm quadrature resonator for transmittance and a 4-element array cryoprobe (Bruker, Switzerland) for signal reception was used. Ultra-fast resting-state fMRI data were acquired with a GE-EPI sequence (TR/TE=90/16ms, FOV=21x21mm2, resolution=250x250um^2, slice thickness=1.2mm, tacq=24min, flip angle=20 degrees, Nreps=16000, 2 scans per animal, BW = 277 kHz axial and coronal). All experiments started at approx. 55 min after bolus injection.
Data analysis:The first 1000 frames were removed due to significant gradient temperature drift during this time, which stabilized after ~800 frames.
Correlation analyses: fMRI signals within the brain mask were bandpass filtered in the conventional rsfMRI range (0.01-0.1Hz) and seed-based correlation maps were performed using Pearson correlation coefficient calculations. Spectral analysis of data: Power spectra were computed voxelwise, after aligning9 and detrending. Spectral power was taken as the integral under the peaks in each bin.
Extracting intrinsic modes from Principle Component Analysis: For each scan, the fMRI signals in N = 1463 brain voxels were band-pass filtered between 0.01 and 0.3Hz and the N × N covariance matrix was computed and averaged across the 20 scans in each group. The first 10 eigen vectors were extracted and mapped.
Results
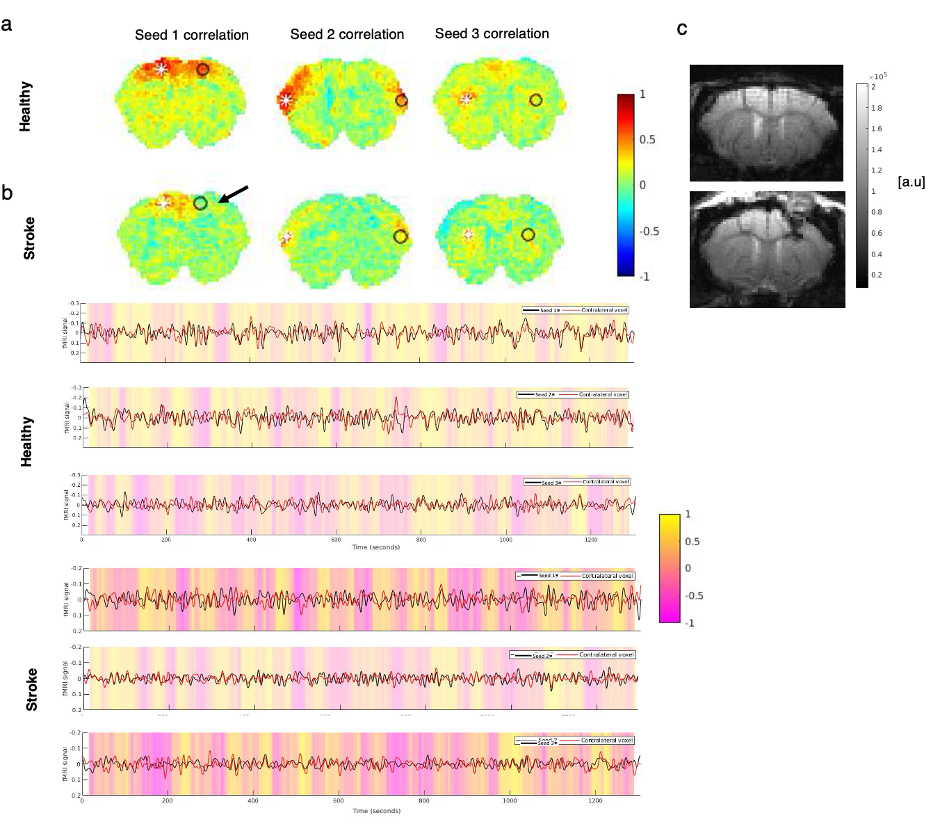
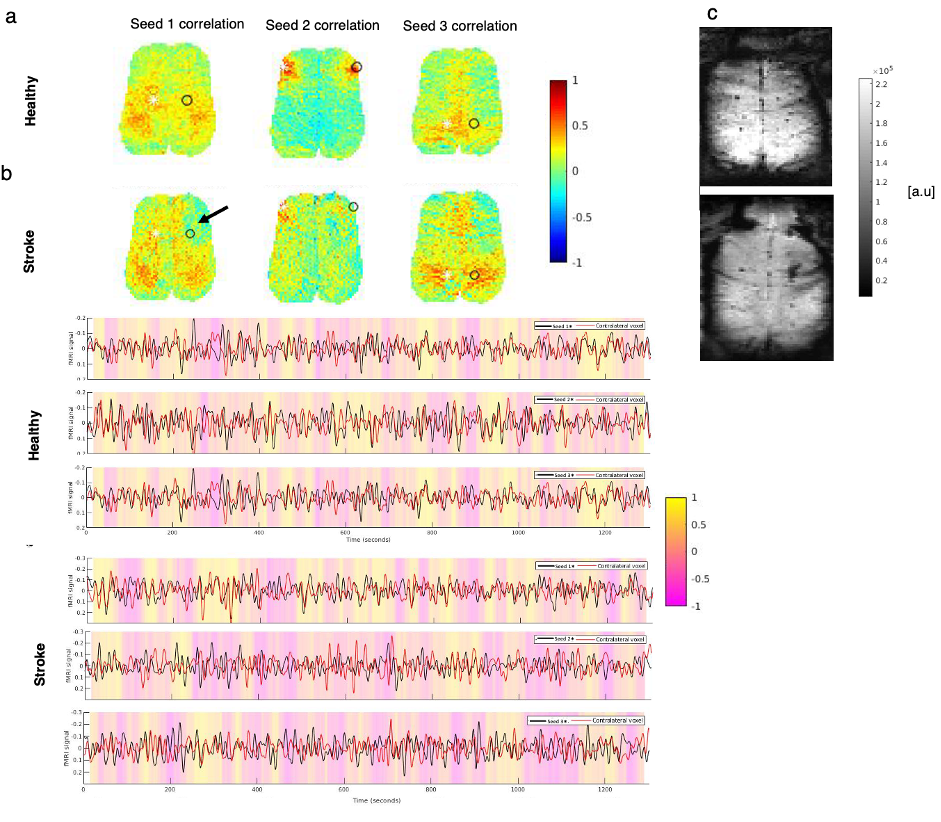
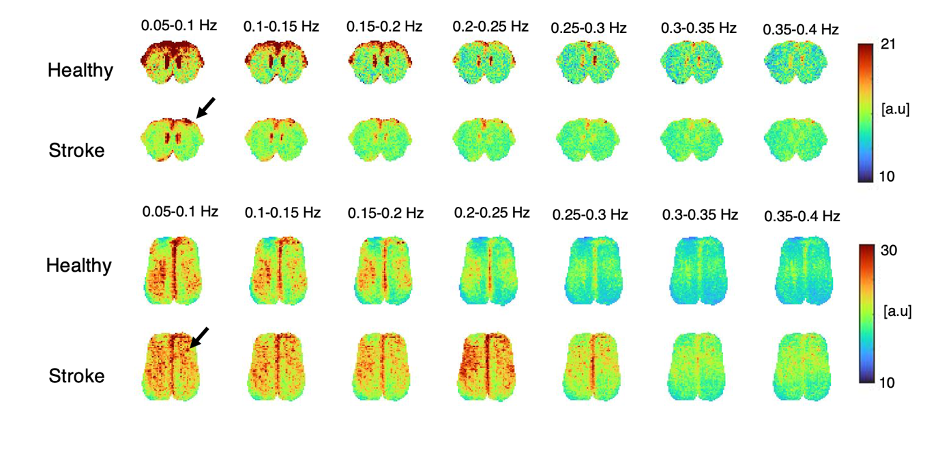
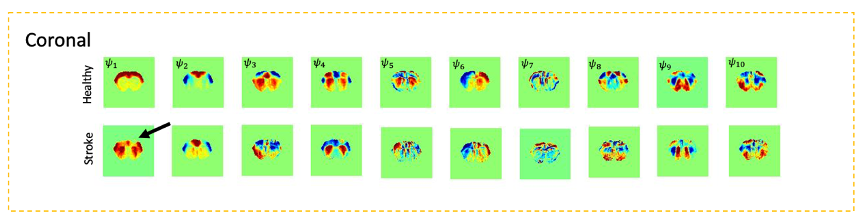
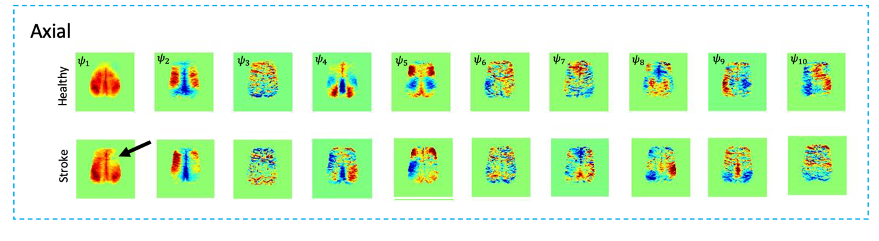
Fig.1 shows a conventional seed-based functional connectivity analysis from the healthy and stroke groups using our ultrafast fMRI data. Long-range correlations were consistently detected across animals, with healthy group seeds showing higher degree of correlation (Fig.1a) compared to the stroke group (Fig. 1b). Particularly, when the contralesional cortex served as a seed, the stroked area showed nearly no connectivity. A more distant contralateral cortical area (seed 2) exhibited similar connectivity to control. Finally, the contralesional striatum (seed 3) showed relatively strong correlation with cortex in the healthy group but less so in stroke. The axial slices revealed similar trends, with strong cortico-cortical correlations in healthy controls both in anterior and posterior areas, but weak correlations near the stroke (Fig.2). Fig. 3 shows the spatial maps of spectral power in 7 non-overlapping frequency bands, averaged across the 20 fMRI scans recorded in each group. Power up to 0.4 Hz is observed in both groups, with clearly higher power in cortex for the healthy group. Fig4 shows the first 10 intrinsic modes detected in both groups for the coronal slice. Modes were similar between individual animals in every group (not shown) but the spatial patterns of these waves were clearly different between the healthy and stroked animals. For instance, φ1 and φ2 show strong cortical oscillations in the healthy controls, while in stroke the oscillations strongly involve striatal areas. Several other modes also show different spatial patterns. Figure 5 shows the first 10 intrinsic modes for the axial slice. Multiple spatial wave patterns are disturbed due to the stroke.Discussion
Our findings likely reflect reorganization of activity in networks in the stroked brain. The weaker FC in the cortico-striatal network is likely due to the conventional filtering below 0.1Hz, while the intrinsic modes revealed strong increases in the cortico-striatal oscillatory patterns. The inter-hemispheric asymmetry observed in both axial and coronal slices in the stroke group may indicate that the contralesional hemisphere is more involved in compensatory mechanisms for the connection breakdown driven by the lesion.Conclusion
The intrinsic mode framework revealed global reorganization following local disruption with enhancement of corticostriatal activity on the expense of the damaged cortico-cortical network. These findings augur well for better understanding of neural network reorganization.Acknowledgements
CONGENTO, PORTUGAL 2020 European Regional Development Fund (ERDF) andFundação para a Ciência e Tecnologia LISBOA-01-0145-FEDER-022170.References
1. Behrens T. E. and Johansen-Berg, H. Relating connectional architecture to grey matter function using diffusion imaging. Philos. Trans. R. Soc. London. B Biol. Sci. 360, 903–911, 2005.
2. Badhwar A., Tam A. et al. Resting state network dysfunction in Alzheimer’s disease: a systematic review and meta-analysis Alzheimers Dement. 8, 73–85, 2017.
3. Bonkhoff A.K, Espinoza F. A, et al. Acute ischemic stroke alters the brain’s preference for distinct dynamic connectivity states. Brain 143, 1525–1540, 2020.
4. van Meer M. P. A., Otte W. M., et al. Extent of bilateral neuronal network reorganization and functional recovery in relation to stroke severity. J. Neurosci. 32, 4495–4507, 2012.
5. van Meer M. P. A van der Marel K. M., et al. Recovery of sensorimotor function after experiemntal stroke correlates with restoration of restingstate interhemispheric functional connectivity. J. Neurosci. 30, 3964–3972, 2010.
6. Pang J, Aquino K, et al. Geometric constraints on human brain function. Nature. 618, 566–574, 2023.
7. Cabral J, Fernandes F.F, Shemesh N. Intrinsic macroscale oscillatory modes driving long range functional connectivity in female rat brains detected by ultrafast fMRI. Nature Communications. 4:375, 2023
8. B. D. Watson, W. D. Dietrich, R. Busto, M. S. Wachtel, and M. D. Ginsberg, “Induction of reproducible brain infarction by photochemically initiated thrombosis,” Ann. Neurol., vol. 17, no. 5, pp. 497–504, 1985.
9. Guizar-sicairos M, Thurman S T, Fienup J R. Efficient subpixel registration. Optics Letters, vol. 33, pp. 156–158, 2008.
Figures