3167
What makes touch social: probing the brain mechanisms supporting affective touch with functional MRI in humans at 3 Tesla1School of Psychology, University of Nottingham, Nottingham, United Kingdom, 2Department of Sports and Exercise Sciences, Manchester Metropolitan University, Manchester, United Kingdom, 3School of Childhood Youth and Education Studies, Manchester Metropolitan University, Manchester, United Kingdom
Synopsis
Keywords: Peripheral Nerves, fMRI (task based), Sensory, Touch, Social
Motivation: Touch is the first sensory modality to develop. C-tactile (CT) nerves in hairy skin are involved in slow/gentle touch, and social interactions. However, brain mechanisms processing the affective aspects of touch remain under-studied.
Goal(s): We characterised the network of brain regions supporting perception of affective touch.
Approach: 11-volunteers underwent fMRI at clinically relevant 3T field strengths while their arm was gently brushed and compared with brushing to remove a marker.
Results: We found a network of brain regions involved in affective touch processing including the posterior superior temporal sulcus (pSTS), inferior frontal gyrus (IFG), temporoparietal junction (TPJ).
Impact: This study characterises the brain network processing affective touch, critical given the central role of touch during development and its social function throughout the lifespan. It helps understanding brain responses in adults born pre-term, where experience of touch was interrupted/reduced.
Introduction
The human skin is the largest and earliest developing sensory system1, with touch having a powerful effect on a child’s growth and development 2–5. In the post-natal period, social touch – positive affective touch – is used between adults and as primary channel of communication between parents and babies6. It is known that C-tactile (CT) nerves in hairy skin are involved in slow, gentle touch, typical of social interactions. However, the role of touch in social communication remains under-studied in basic neuroscience, especially compared to other sensory modalities, specifically vision and hearing7. The brain responses to touch are still poorly understood. This project uses functional Magnetic Resonance Imaging at clinically relevant field strengths (3 Tesla) to understand brain responses to social touch in adults. We address the question, do brain responses to touch differ depending on the perceived goal of the tactile experience?Methods
All data was acquired on a Siemens 3T MAGNETOM Vida system (preinstalled XQ gradients 45mT/m @ 200 T/m/s), using a 64-channel head/neck coil at the Institute of Sport, following approval by the Science and Engineering ethics review board at Manchester Metropolitan University (Manchester, UK). Functional scans were performed (N=11) using a whole brain 2D GRE-EPI sequence, with voxel size 2mm*2mm*3.75mm (28 slices, TR/TE=1000/23ms, pF=6/8, flip angle=70º, Multi-Band factor=4, bandwidth=1834 Hz/Px, FoV=191mm). One participant was omitted from analysis due to the known effects with confounding neurodivergence8. Whole-brain T1-weighted structural images (0.8mm isotropic) were acquired using standard MP2RAGE.Paradigm: Participants performed between 2 and 8 functional runs (~6min each) per imaging session. Functional scans consisted of initial 60s of fixation, followed by six 25s ON-OFF blocks where either the right or left (counterbalanced) arm was gently brushed using a smooth xylophone wooden mallet (thin stick and smooth rounded head; figure 1). In the social condition, participants simply received the gentle brushing on their arm. In the goal condition, the experimenter performed the brushing as a mean to remove a pen marker on the participant’s arm. CTs are known to be velocity tuned and brushing speed used in the present study was informed by McGlone et.al. (2014)9.
Data analysis: Structural data was automatically segmented using Freesurfer10. Functional data was analysed using FSL FEAT11. Preprocessing included slice time correction and high-pass filtering, motion correction (MCFLIRT) and co-registration to within session anatomical images using boundary based registration. A general linear model (GLM) was applied to obtain an estimate of the response to touch (social/goal) against baseline. Functional datasets were then normalised to MNI space to perform group-level analysis and determine the network of brain areas involved in processing social/goal-directed touch.
Results & Discussion
To determine the brain mechanisms supporting affective touch we obtained statistical maps (following GLM-based analysis at a group level, with cluster-threshold of z>3.1 and correction for multiple comparisons corresponding to α<0.05) to each touch condition (social and goal) relative to baseline. The results (figure 2) revealed robust activation in the right somatosensory cortex (pre and post central sulcus), temporoparietal junction, inferior frontal gyrus. Interestingly, activations in posterior superior temporal sulcus, supramarginal gyrus, and posterior insula were more pronounced in the social compared to goal condition (figure 2b). The haemodynamic response time course within the posterior superior temporal sulcus (pSTS) is markedly different across the two touch conditions (figure 3). These results highlight the involvement of brain regions such as the pSTS in processing not only social elements through vision and sound, but also touch.Conclusion & Summary
Warm physical contact and affective touch are critical parts of the earliest interactions between parents and infants, and fundamental throughout the lifespan12. Characterising the brain mechanisms underpinning affective touch provides insights in disorders where touch dysfunction is observed13 (e.g., Autism Spectrum Disorder), and presents a stepping stone in understanding these mechanisms in adults born pre-term, where experience of touch was interrupted (pre-term birth) or reduced (low skin-to-skin tactile stimulation). Seemingly, recent studies14 suggest that gentle skin stroking, activating CT-fibres, can help reduce perceived pain caused by high pressure applied to the thumb. Understanding affective touch mechanisms could therefore pave the way for non-pharmacological analgesic solutions.Acknowledgements
AJK/JMS/LS acknowledge provision of MR scanning time from the Manchester Metropolitan University.References
1. Field, T. Touch for socioemotional and physical well-being: A review. Dev. Rev. 30, 367–383 (2010).
2. Cascio, C. J., Moore, D. & McGlone, F. Social touch and human development. Dev. Cogn. Neurosci. 35, 5–11 (2019).
3. Feldman, R., Rosenthal, Z. & Eidelman, A. I. Maternal-preterm skin-to-skin contact enhances child physiologic organization and cognitive control across the first 10 years of life. Biol. Psychiatry 75, 56–64 (2014).
4. Mantis, I., Mercuri, M., Stack, D. M. & Field, T. M. Depressed and non-depressed mothers’ touching during social interactions with their infants. Dev. Cogn. Neurosci. 35, 57–65 (2019).
5. Rigato, S. et al. Cortical signatures of vicarious tactile experience in four-month-old infants. Dev. Cogn. Neurosci. 35, 75–80 (2019).
6. Sailer, U. & Leknes, S. Meaning makes touch affective. Curr. Opin. Behav. Sci. 44, 101099 (2022).
7. Gallace, A. & Spence, C. The science of interpersonal touch: an overview. Neurosci. Biobehav. Rev. 34, 246–259 (2010).
8. Kaiser, M. D. et al. Brain mechanisms for processing affective (and nonaffective) touch are atypical in autism. Cereb. Cortex 26, 2705–2714 (2016).
9. McGlone, F., Wessberg, J. & Olausson, H. Discriminative and affective touch: sensing and feeling. Neuron 82, 737–755 (2014).
10. FreeSurfer. FreeSurfer http://surfer.nmr.mgh.harvard.edu/.
11. Jezzard, P., Matthews, P. M. & Smith, S. M. Functional MRI. (Oxford University Press, 2001).
12. Essick, G. K. et al. Quantitative assessment of pleasant touch. Neurosci. Biobehav. Rev. 34, 192–203 (2010).
13. Cascio, C. J. Somatosensory processing in neurodevelopmental disorders. J. Neurodev. Disord. 2, 62–69 (2010).
14. Savallampi, M. et al. Social touch reduces pain perception-an fMRI study of cortical mechanisms. Brain Sci. 13, (2023).
Figures
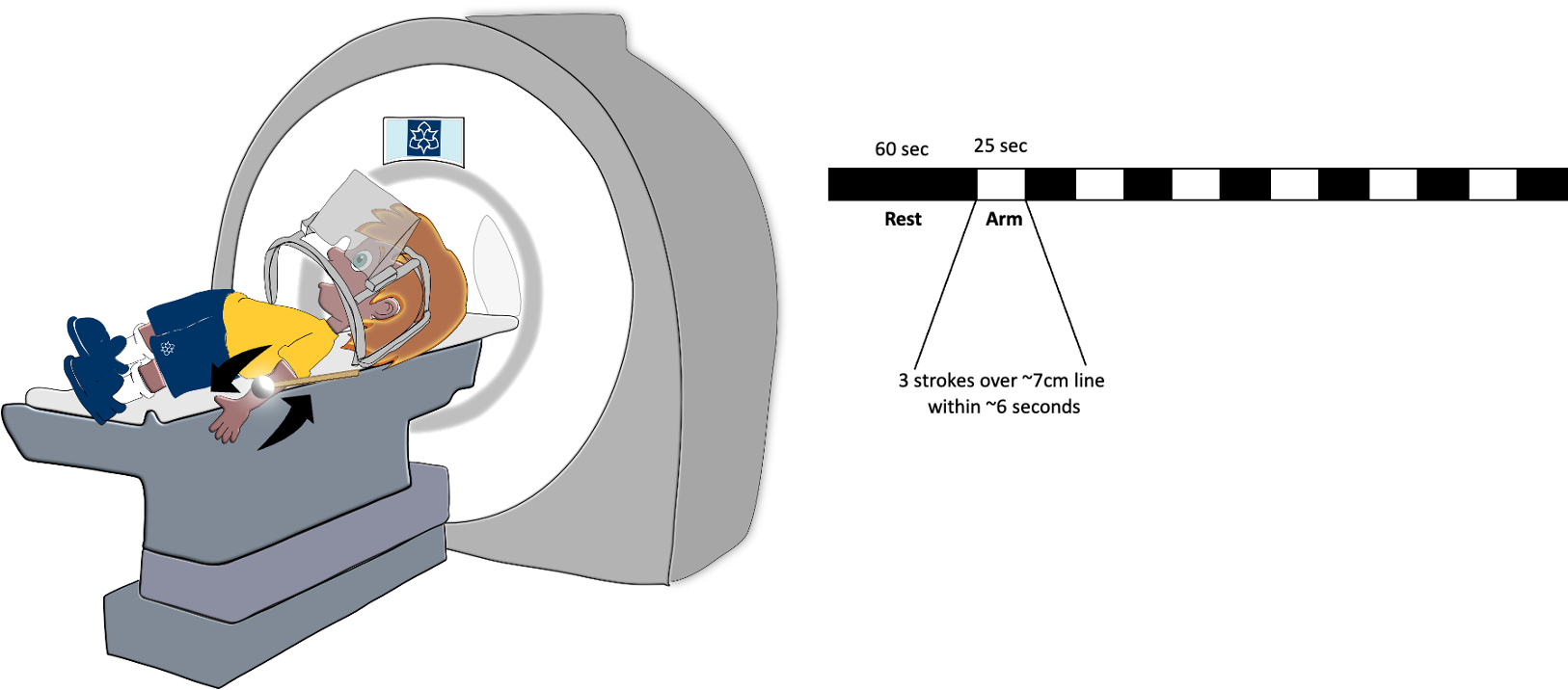
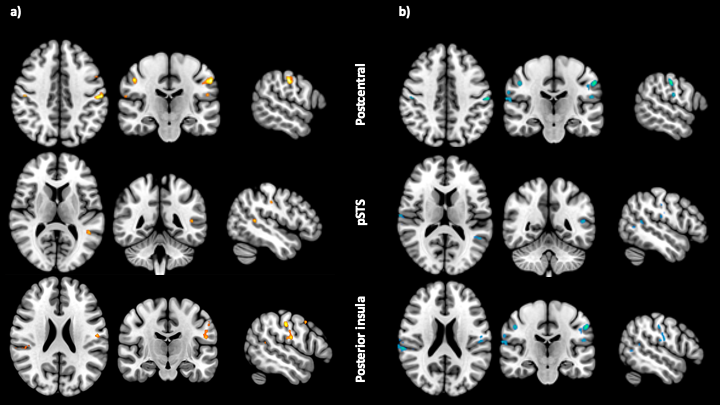
Figure 2 - Group level analysis results. a) Statistical maps identifying clusters in the goal vs baseline condition. Thresholded clusters show postcentral sulcus, posterior superior temporal sulcus, and posterior insula gyrus activation. b) Statistical maps identifying clusters of activity in the social vs baseline condition. Same areas as in a) for comparison purposes.
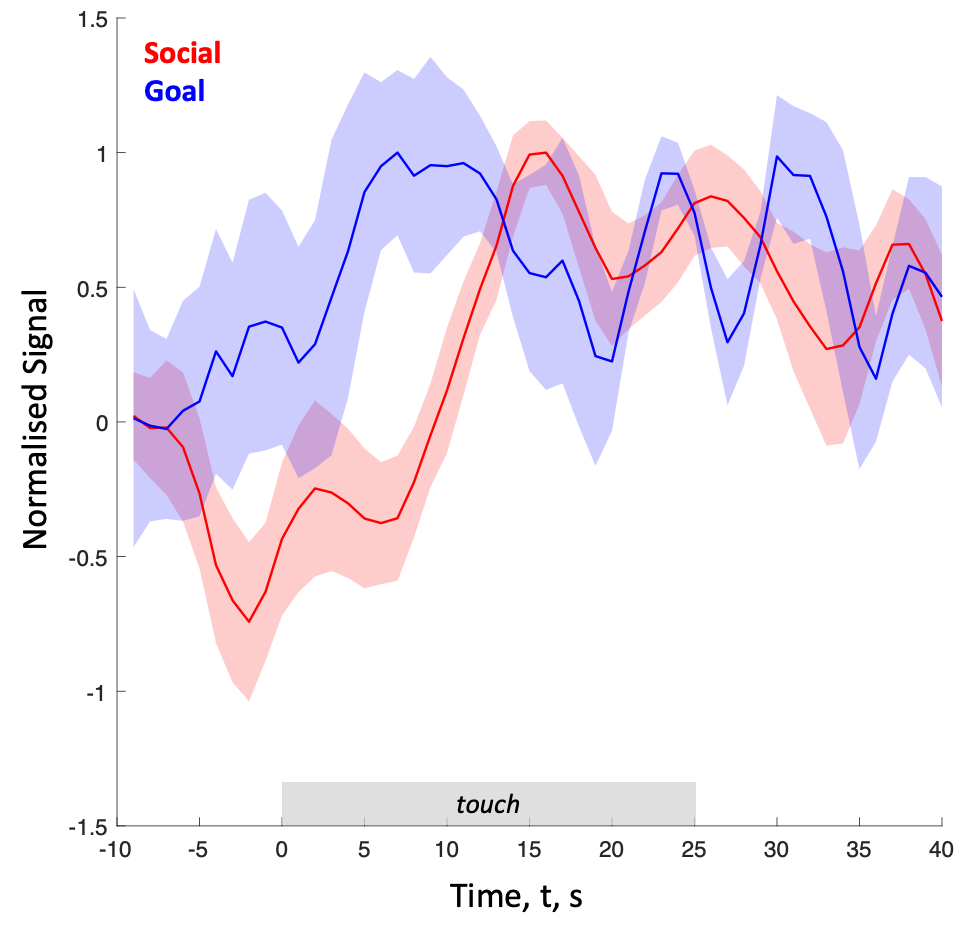
Figure 3 - Average time course of the normalised BOLD response from the voxels in the right posterior superior temporal sulcus (pSTS) for the social (red) and goal (blue) conditions. Shaded area shows standard errors of the means. The 0s time point represents the onset of the brushing (touch).