3126
Small voxel sizes reduce extravascular dephasing from large veins in gradient-echo BOLD fMRI at 7T: a simulation study1Department of Physics, Carleton University, Ottawa, ON, Canada, 2University of Ottawa Institute of Mental Health Research, Royal Ottawa Mental Health Centre, Ottawa, ON, Canada, 3Department of Radiology, Boston Children's Hospital, Boston, MA, United States, 4Department of Radiology, Harvard Medical School, Boston, MA, United States, 5Department of Radiology, Stanford University, Stanford, CA, United States, 6Department of Electrical Engineering, Stanford University, Stanford, CA, United States, 7Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital, Charlestown, MA, United States, 8Division of Health Sciences and Technology, Massachusetts Institute of Technology, Cambridge, MA, United States
Synopsis
Keywords: fMRI Acquisition, High-Field MRI, BOLD fMRI; Biophysical Simulations; High-Resolution fMRI; Vasculature
Motivation: Recent ultra-high-resolution gradient-echo BOLD-fMRI data demonstrate an unexpected reduction in sensitivity to large veins at 7T, suggesting that small voxels may experience less signal loss due to extravascular dephasing.
Goal(s): To test whether a reduction in through-plane or in-plane dephasing with small voxels may measurably contribute to reduced large-vein influences in gradient-echo BOLD.
Approach: We apply biophysical simulations of simplified vascular architecture, combined with realistic image encoding gradients.
Results: While voxels adjacent to veins are still strongly influenced, smaller voxel sizes have reduced extravascular BOLD, which impacts cerebral cortical-depth profiles of activation. The direction of the imaging encoding gradients also affects these profiles.
Impact: Rencelty available ultra-high-resolution fMRI improves spatial accuracy, and may also provide an unexpected improvement in neuronal specificity. We demonstrate that small-voxel gradient-echo BOLD achieves reduced unwanted contamination from large veins, potentially providing an fMRI method with high sensitivity AND specificity.
Introduction
Advances in ultra-high field MRI have enabled small fMRI voxel sizes at the ~500-micron scale with sufficient sensitivity to detect neuronally-driven activation1,2. While small voxels provide increased imaging resolution and reduced contributions of physiological noise and related instabilities3–5, they may also confer increased microvascular specificity in gradient-echo BOLD. Recent observations based on 400-micron gradient-echo BOLD fMRI data suggest that unwanted large-vein effects may be reduced with smaller gradient-echo EPI voxel sizes6. Intuitively, extravascular dephasing may be reduced with smaller voxels—akin to the well-known effect of reducing through-plane dephasing and increasing T2* by adopting thinner slices in 2D gradient-echo imaging—which would lead to reduced large-vein contributions at ultra-high fields. This effect of voxel size on signal loss in the presence of static field inhomogeneity has been considered previously7–9, and it was found that losses due to dephasing from strong field gradients near, e.g., air-tissue interfaces can be restored by increasing resolution in 2D and 3D imaging.To characterize how extravascular dephasing from large veins is reduced with smaller voxel sizes in gradient-echo BOLD, we performed a series of biophysical simulations using simplified models of venous vasculature and realistic imaging gradients.
Methods
We simulated the gradient-echo BOLD signal at 7T from a 2D voxel located next to a large pial vein perpendicular to B0 (Figure 1A) using an echo-time of 27ms. We simulated the BOLD signal from the vein alone as the distance of the voxel from the pial surface was increased and, similarly, from a voxel containing only parenchymal capillaries of 4-μm radius. Simulations used voxels of size 0.5–1.0 mm and were repeated without and with image-encoding effects. Simulations that ignored image-encoding were representative of through-plane gradient effects. Only extravascular signal was considered, however, the results for the total BOLD signal were not significantly different given the short T2* of veins10.We applied our previous framework accounting for the effects of image-encoding and EPI readout duration11. Figure 1B shows a schematic of the imaging slice. EPI phase-encoding (PE) gradients with a range of echo-spacings, directions, and polarities were considered (Figure 1C) while frequency-encoding was ignored. We also retrospectively modelled image-encoding by scaling simulations without PE gradients by the desired k-space weighting at each echo of the echo-train (Figure 1D).
To quantify the influence of pial veins, we computed the “critical depth” as the distance from the pial surface at which the BOLD signal from capillaries surpasses that of the pial vein12.
Results
Figure 2 demonstrates the cortical depth-dependence of the pial vein extravascular dephasing for multiple pial vein radii and voxel sizes, ignoring image-encoding. The depth-dependence is examined using baseline R2', change in R2' during activation, and the percent signal change. In all cases, the smallest voxel size (0.5 mm) has the most dephasing at the surface, but its dephasing decreases most rapidly with depth.Examples of the impact of image-encoding on the depth-dependence are shown in Figure 3 for a 32-ms acquisition window. Depending on the PE direction and polarity, the BOLD signal amplitude from the pial vein may be increased or decreased, particularly when the gradient is along the radial direction. Retrospective phase-encoding can capture the average effect of the explicit encoding gradients and is comparable to the results using tangentially-oriented PE gradients. The BOLD signal at two depths is summarized in Figure 4, and the critical depth when the capillary BOLD signal dominates is given in Figure 5. While the depth at which the pial vein dominates shifts closer to the pial surface with smaller voxels, the BOLD signal change within those small voxels at the pial surface becomes larger, indicating that the large-vein BOLD contributions become more concentrated at the pial surface with smaller voxels. Longer EPI readouts may also amplify dephasing from the larger pial veins.
Discussion and Conclusion
While both through-plane and in-plane dephasing contribute to signal loss in gradient-echo imaging, in-plane dephasing will depend on the direction and magnitude of the image encoding gradients. Because of the complex folding pattern of the human cerebral cortex, this implies that the effects of pial veins will vary across cortical locations.Our findings suggest that small-voxel gradient-echo BOLD may be capable of enhanced microvascular specificity, which may make BOLD a more attractive fMRI contrast, provided one can achieve sufficiently small voxels. Similarly, non-BOLD fMRI methods with gradient-echo EPI readouts13 may also benefit from having less large-vein BOLD contamination to correct. To increase SNR without reintroducing large-vein contamination, anatomically-informed smoothing of the magnitude images could be employed4. Future work will consider dephasing effects on anatomically realistic vascular models14,15.
Acknowledgements
We would like to thank Drs. Renzo Huber and David Feinberg for helpful discussions. This work was supported in part by the NIH NIBIB (grants P41-EB030006, R01-EB019437, R01-EB033206 and R01-EB032746), by the BRAIN Initiative (NIH NIMH grant R01-MH111419 and NIH NINDS grant U19-NS123717), by the MGH/HST Athinoula A. Martinos Center for Biomedical Imaging, and by Carleton University.References
1. Berman AJL, Grissom WA, Witzel T, et al. Ultra-high spatial resolution BOLD fMRI in humans using combined segmented-accelerated VFA-FLEET with a recursive RF pulse design. Magn Reson Med. 2021;85(1):120-139. doi:10.1002/mrm.28415
2. Feinberg DA, Beckett A, Vu JA, et al. Next Generation MRI scanner designed for ultra-high resolution human brain imaging at 7-Tesla. Nat Methods.
3. Huber L (Renzo), Ivanov D, Handwerker DA, et al. Techniques for blood volume fMRI with VASO: From low-resolution mapping towards sub-millimeter layer-dependent applications. Neuroimage. 2018;164:131-143. doi:10.1016/j.neuroimage.2016.11.039
4. Błażejewska AI, Fischl B, Wald LL, Polimeni JR. Intracortical smoothing of small-voxel fMRI data can provide increased detection power without spatial resolution losses compared to conventional large-voxel fMRI data. Neuroimage. 2019;189:601-614. doi:10.1016/j.neuroimage.2019.01.054
5. Triantafyllou C, Hoge RD, Wald LL. Effect of spatial smoothing on physiological noise in high-resolution fMRI. Neuroimage. 2006;32(2):551-557. doi:10.1016/j.neuroimage.2006.04.182
6. Feinberg DA, Torrisi S, Beckett AJ, et al. Sub-0.1 microliter CBV fMRI on the Next Generation 7T scanner. Proc Intl Soc Mag Reson Med. 2022;30:0397. https://submissions.mirasmart.com/ISMRM2022/Itinerary/Files/PDFFiles/0397.html.
7. Reichenbach JR, Venkatesan R, Yablonskiy DA, Thompson MR, Lai S, Haacke EM. Theory and application of static field inhomogeneity effects in gradient-echo imaging. Journal of Magnetic Resonance Imaging. 1997;7(2):266-279. doi:10.1002/JMRI.1880070203
8. Haacke EM, Tkach JA, Parrish TB. Reduction of T2* dephasing in gradient field-echo imaging. Radiology. 1989;170(2):457-462. doi:10.1148/RADIOLOGY.170.2.2911669
9. Hernando D, Vigen KK, Shimakawa A, Reeder SB. R2* mapping in the presence of macroscopic B0 field variations. Magn Reson Med. 2012;68(3):830-840. doi:10.1002/MRM.23306
10. Uludağ K, Müller-Bierl B, Uğurbil K. An integrative model for neuronal activity-induced signal changes for gradient and spin echo functional imaging. Neuroimage. 2009;48(1):150-165. doi:10.1016/j.neuroimage.2009.05.051
11. Berman AJL, Wang F, Setsompop K, Chen JJ, Polimeni JR. Biophysical simulations of the BOLD fMRI signal using realistic imaging gradients: Understanding macrovascular contamination in Spin-Echo EPI. Proc Intl Soc Mag Reson Med. 2021;29:3398. https://index.mirasmart.com/ISMRM2021/PDFfiles/3398.html.
12. Pfannmoeller JP, Berman AJL, Kura S, Cheng X, Boas DA, Polimeni JR. Quantification of draining vein dominance across cortical depths in BOLD fMRI from first principles using realistic Vascular Anatomical Networks. Proc Intl Soc Mag Reson Med. 2019;27:3715.
13. Huber L (Renzo), Ivanov D, Krieger SN, et al. Slab-selective, BOLD-corrected VASO at 7 Tesla provides measures of cerebral blood volume reactivity with high signal-to-noise ratio. Magn Reson Med. 2014;72(1):137-148. doi:10.1002/mrm.24916
14. Gagnon L, Sakadžić S, Lesage F, et al. Quantifying the microvascular origin of bold-fMRI from first principles with two-photon microscopy and an oxygen-sensitive nanoprobe. Journal of Neuroscience. 2015;35(8):3663-3675. doi:10.1523/JNEUROSCI.3555-14.2015
15. Hartung G, Pfannmoeller JP, Berman AJL, Polimeni JR. Simulated fMRI responses using human Vascular Anatomical Network models with varying architecture and dynamics. Proc Intl Soc Mag Reson Med. 2022;30:0682. https://submissions.mirasmart.com/ISMRM2022/Itinerary/Files/PDFFiles/0682.html.Figures
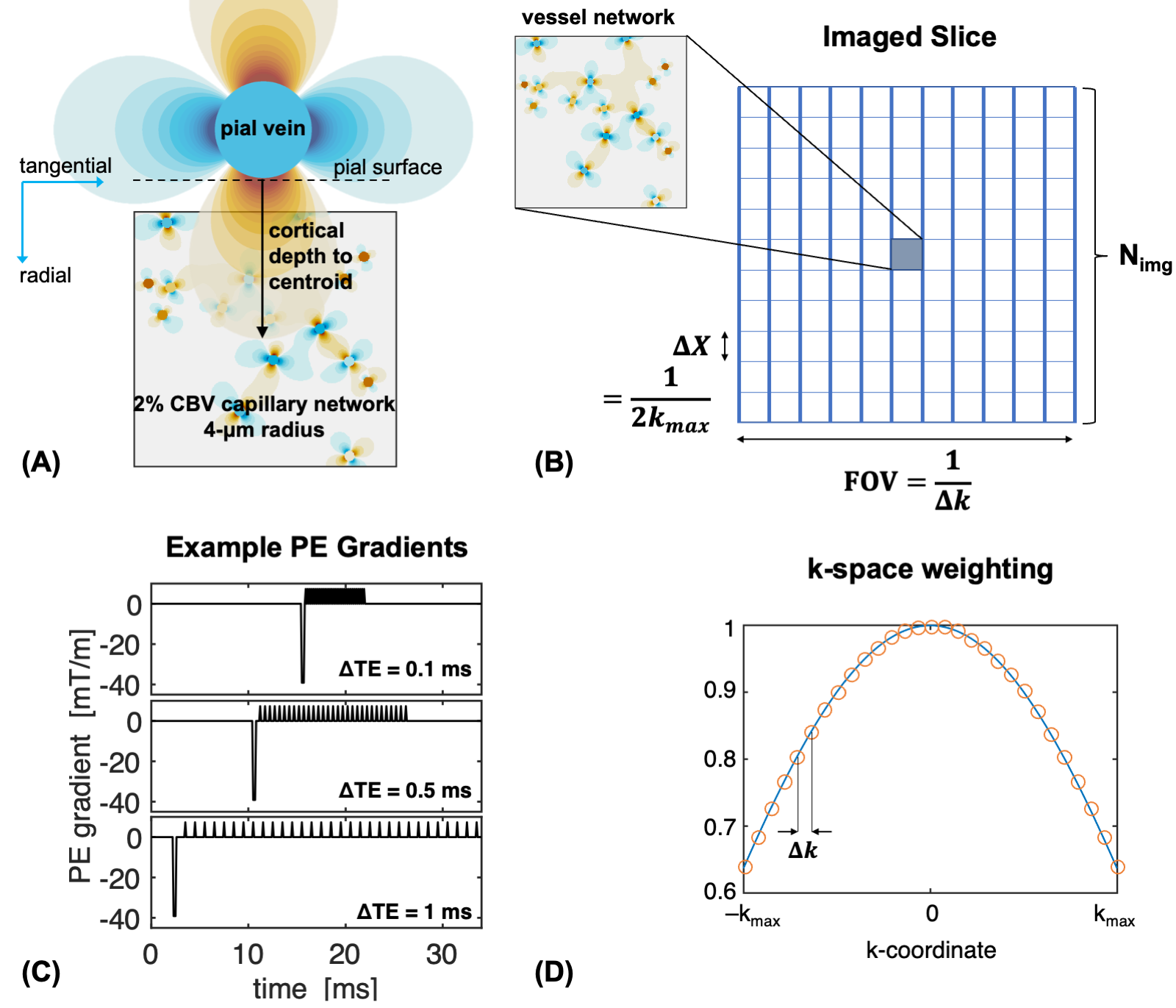
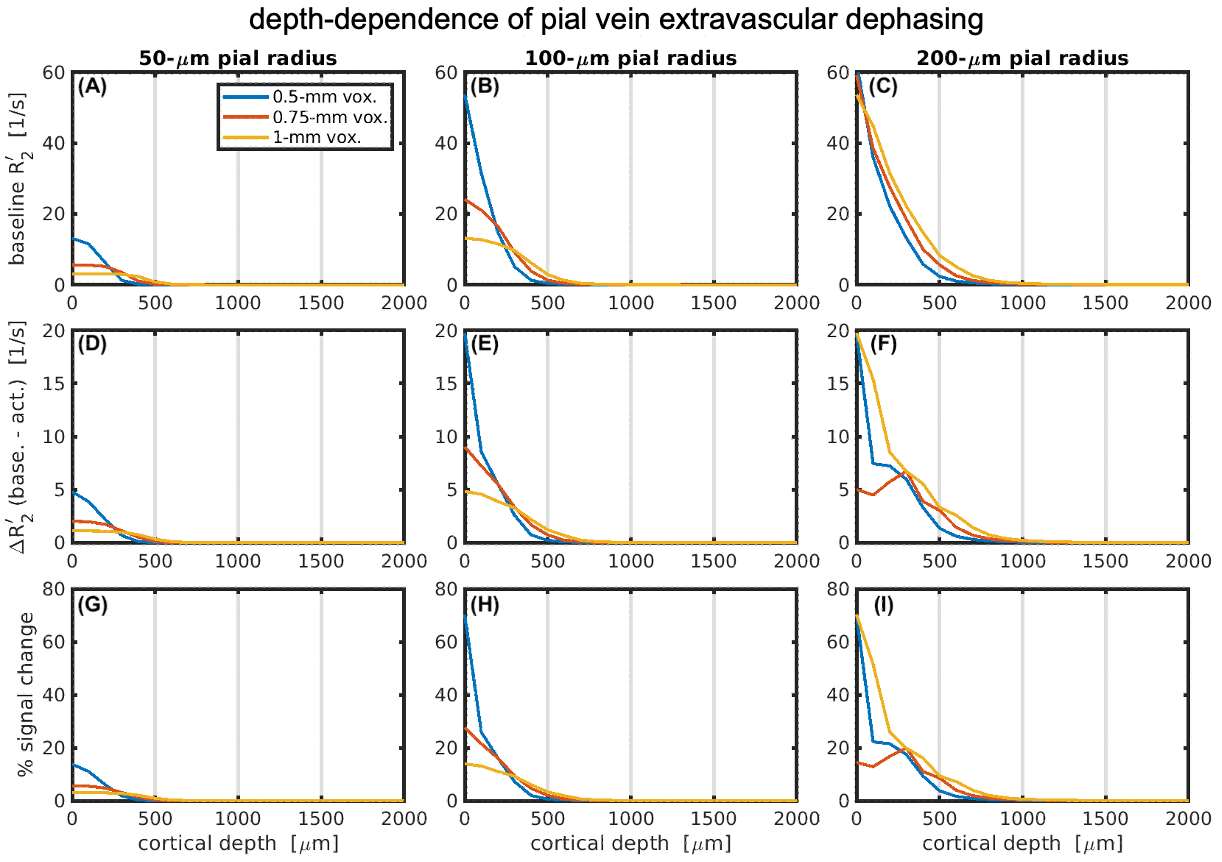
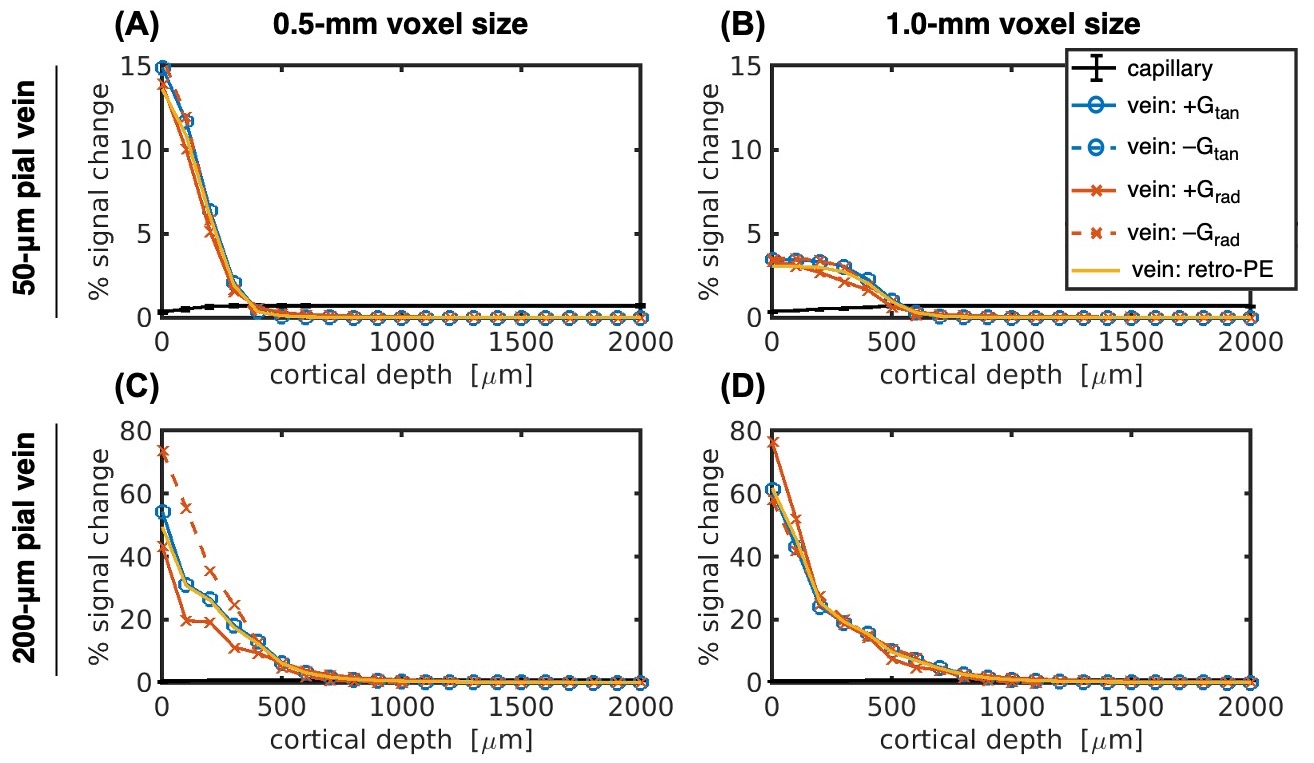
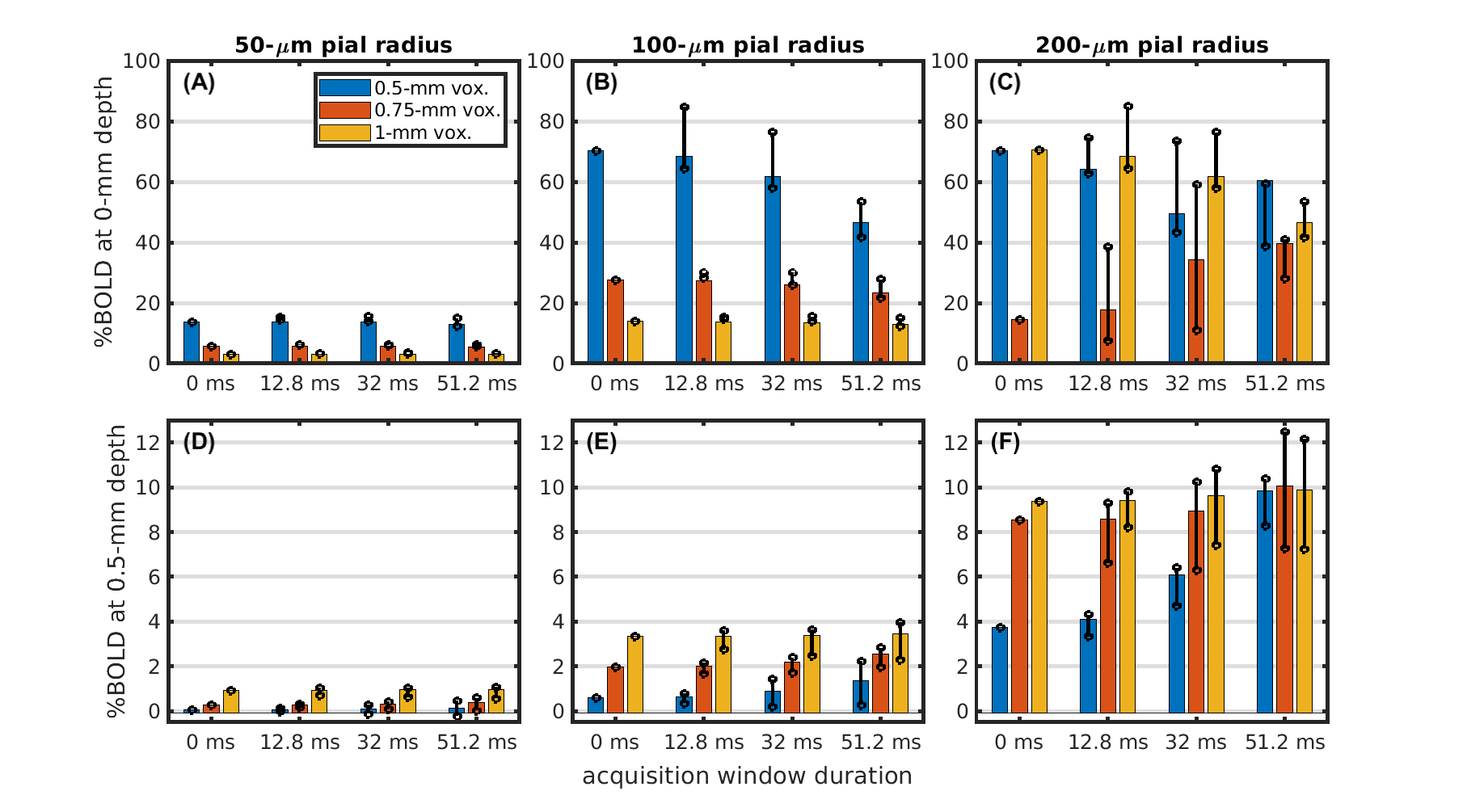
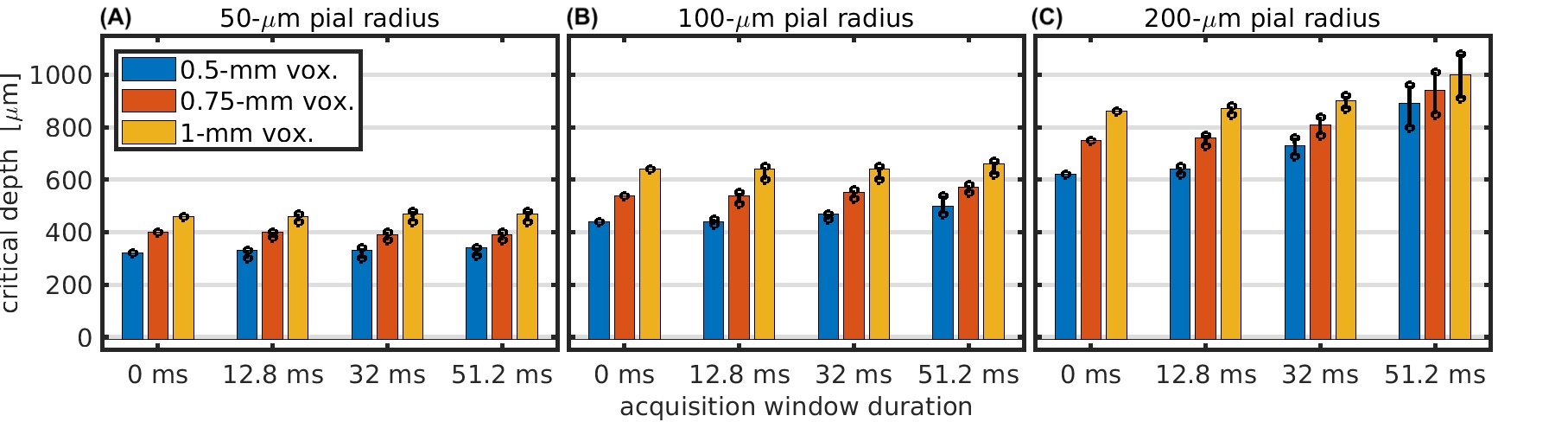
Fig 5: Critical depth where the BOLD signal of the pial vein drops below that of the capillary network. Critical depth is shown vs. acquisition window duration for pial vein radii of 50 μm (A), 100 μm (B), and 200 μm (C) and for voxel sizes of 0.5 mm (blue), 0.75 mm (orange), and 1 mm (yellow). The bar amplitude is the critical depth when using retrospective phase-encoding. The circle markers represent the minimum/maximum critical depth when using the explicit gradients for phase-encoding.