2517
Investigating brain correlates of neural plasticity with visual-motor sequence learning: Preliminary observations using 3.0T MAGNUS1Technology & Innovation Center, GE HealthCare, Niskayuna, NY, United States, 2Concordia University, Montreal, QC, Canada, 3GE HealthCare, Mississauga, ON, Canada
Synopsis
Keywords: White Matter, Brain Connectivity, High performance gradient MRI, WM plasticity, Learning, Dynamic white matter plasticity
Motivation: To investigated whether white matter microstructural changes after adaptive short-term training can be detected in a limited sample size with a high-performance gradient system that allows for increased sensitivity to underlying brain micro-architecture.
Goal(s): Demonstrate that with task complexity and short-term training, the dynamic-location-specific neuroplastic changes can be detected.
Approach: Advanced structural and functional MRI sequences in <60-min examination with a pre- and post-design paradigm were used in a high-performance head gradient system that allows for increased sensitivity to the underlying brain micro-architecture.
Results: In this pilot study microstructural changes were noted on a group basis underlying primary motor and sensorimotor cortices.
Impact: This study highlights the role of dynamic brain processes following short term visual motor sequencing learning (VMSL). The adult brain preserves dynamic characteristics that are altered by short-term learning experiences that can be detected with dMRI.
Introduction
Neuroplasticity is fundamentally linked to learning and retention, brain repair and reorganization, and may be impacted in neurodegeneration. The underlying plastic processes are highly clinically relevant particularly if changes in brain microarchitecture can be assessed non-invasively. A growing number of studies have highlighted the plastic potential of WM, with alterations reported not only as a consequence of long-term potentiation, but also with short-term learning/training paradigms[1]. WM changes based on DTI have been reported across various temporal scales ranging from hours to weeks of training. Sagi, et al. showed that 2h of video game training induced decrease in MD[2], with others reporting detection of plastic molding over weeks of juggling[3] and verbal and spatial span tasks[4,5].The study of brain plasticity using MRI has historically tended to focus on T1w and tensor metrics potentially due to performance limitations of conventional clinical scanners, and the limits to sensitivity of metrics derived from tensor fitting, which though sensitive to the tissue microenvironment, are non-specific to the type of change. With the capabilities of high-performance gradient systems[6-8] these metrics can be evaluated in a more relevant parameter space (shorter TEs, pulse-widths), while ultra-high b-value (≥10-30ms/μm2) diffusion encoding can be leveraged to explore simplified models of intra-axonal diffusivity. Furthermore, shorter echo-spacing allows multi-echo fMRI to be acquired without loss in temporal resolution, providing enhanced BOLD sensitivity with information of dynamic T2* changes that better reflect neuronal activity[9]. The gradient performance advantages translate such that in a 60-min scan, a comprehensive parameter space can be explored, where a combination of contrasts offers a complementary but powerful mechanism to understand how experiences shape the brain.
In this preliminary study, we leveraged a well-described activity-dependent short-term training paradigm with reported potentiation that tracks learning stages and behavioral outcomes: Visual Motor Sequence Learning(VMSL). A comprehensive acquisition space was explored with MAGNUS for assessing brain microarchitecture changes in response to pinch-force VMSL in pre-post design paradigm setting.
Methods
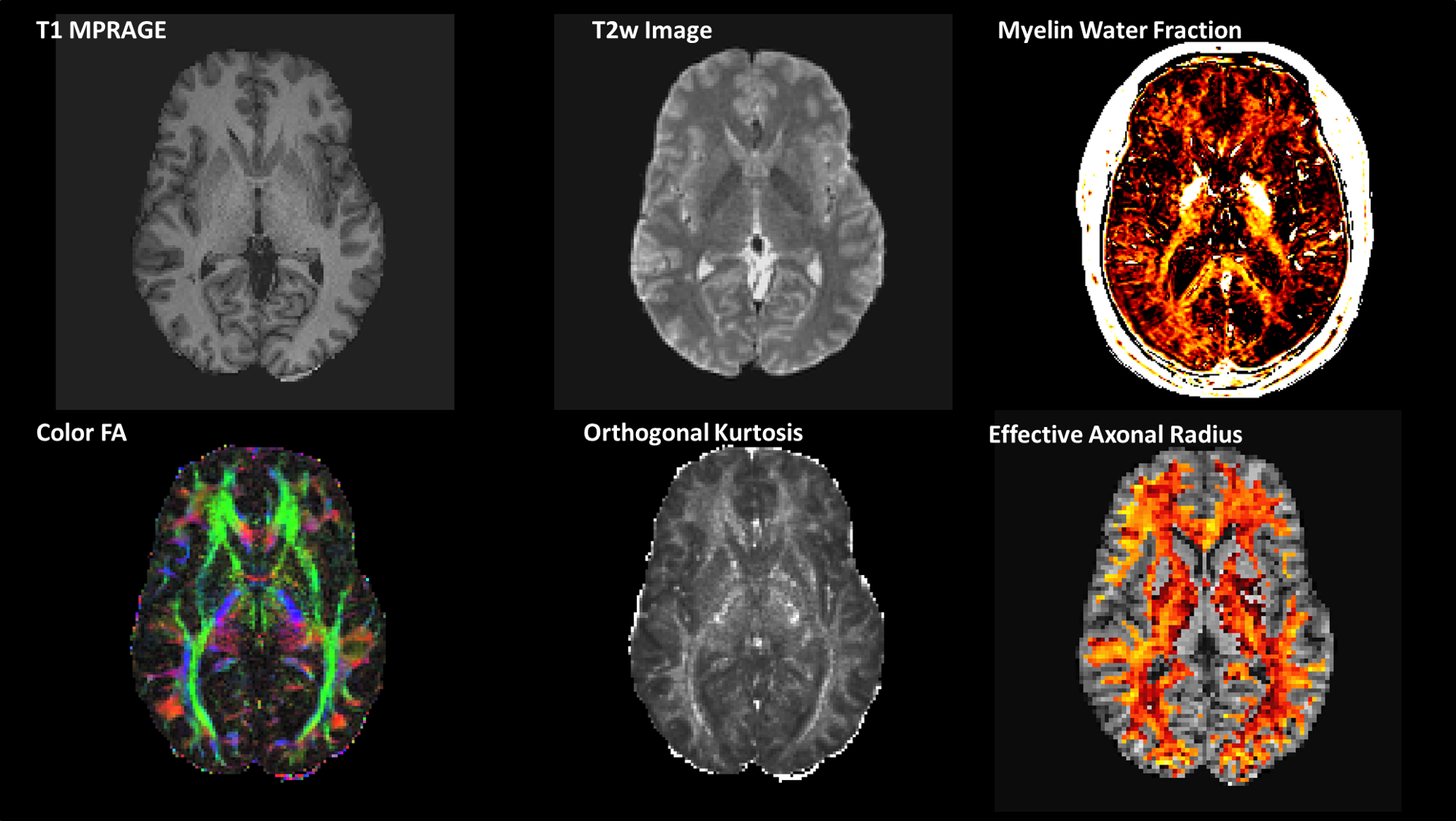
Acquisition: Five (Male, age = 46 ± 11.4 yrs) volunteers were recruited and scanned with identical protocols on two consecutive days with the MAGNUS[6] (Gmax,SRmax =300mT/m,750T/m/s) gradient, under IRB-approved protocols. A 32-channel phased array head coil (NOVA Medical, Wilmington, MA, USA) was used for all scans. The scan protocol is summarized in Table 1. A pinch force VMSL task was administered outside of the scanner following each imaging session. Participants were tasked with matching the position of a force bar with a reference bar that moved based on a pre-determined complex sequence. The sequence was performed across 9 blocks (three trials/block), with a rest period of the same duration between each.Signal processing: Diffusion-weighted images were corrected for eddy current distortion, bulk motion, susceptibility and gradient non-linearity correction[10], followed by generalized spherical deconvolution for denoising[11], using a custom image processing pipeline. Myelin water fraction was computed by fitting a multi-exponential T2 decay curve using DECAES[12]. Ultra-High-b Real-valued data[13] were processed with the same custom processing pipeline. Spherical mean signal was modeled to generate a projection of the tail-weighted reff (µm) distribution[14].
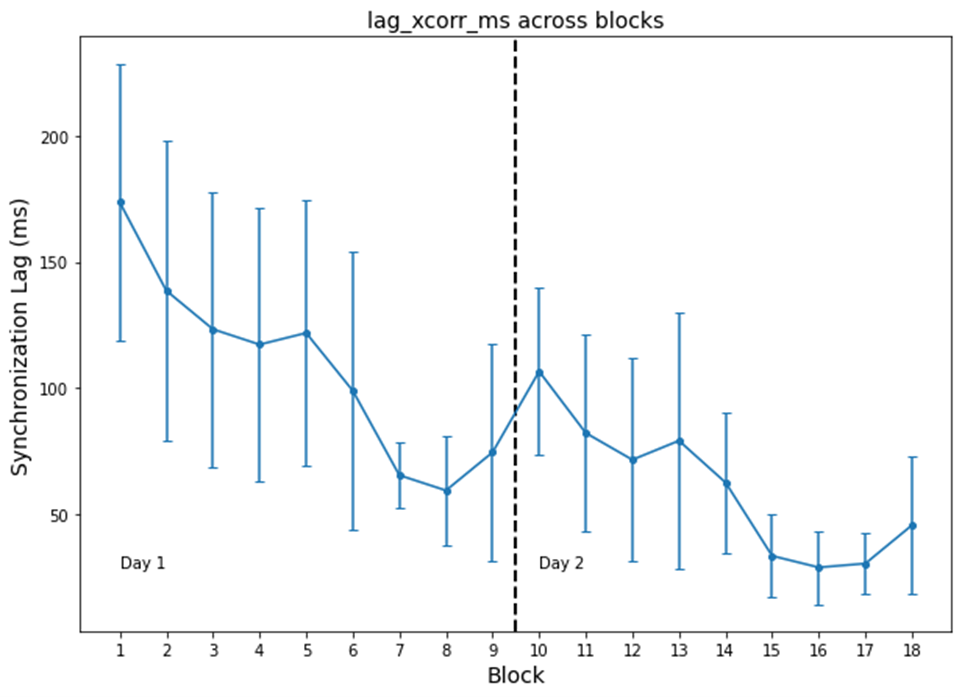
VMSL: Performance was assessed with measures of synchrony (temporal lag) and root mean squared error (height deviation) across learning and between days.
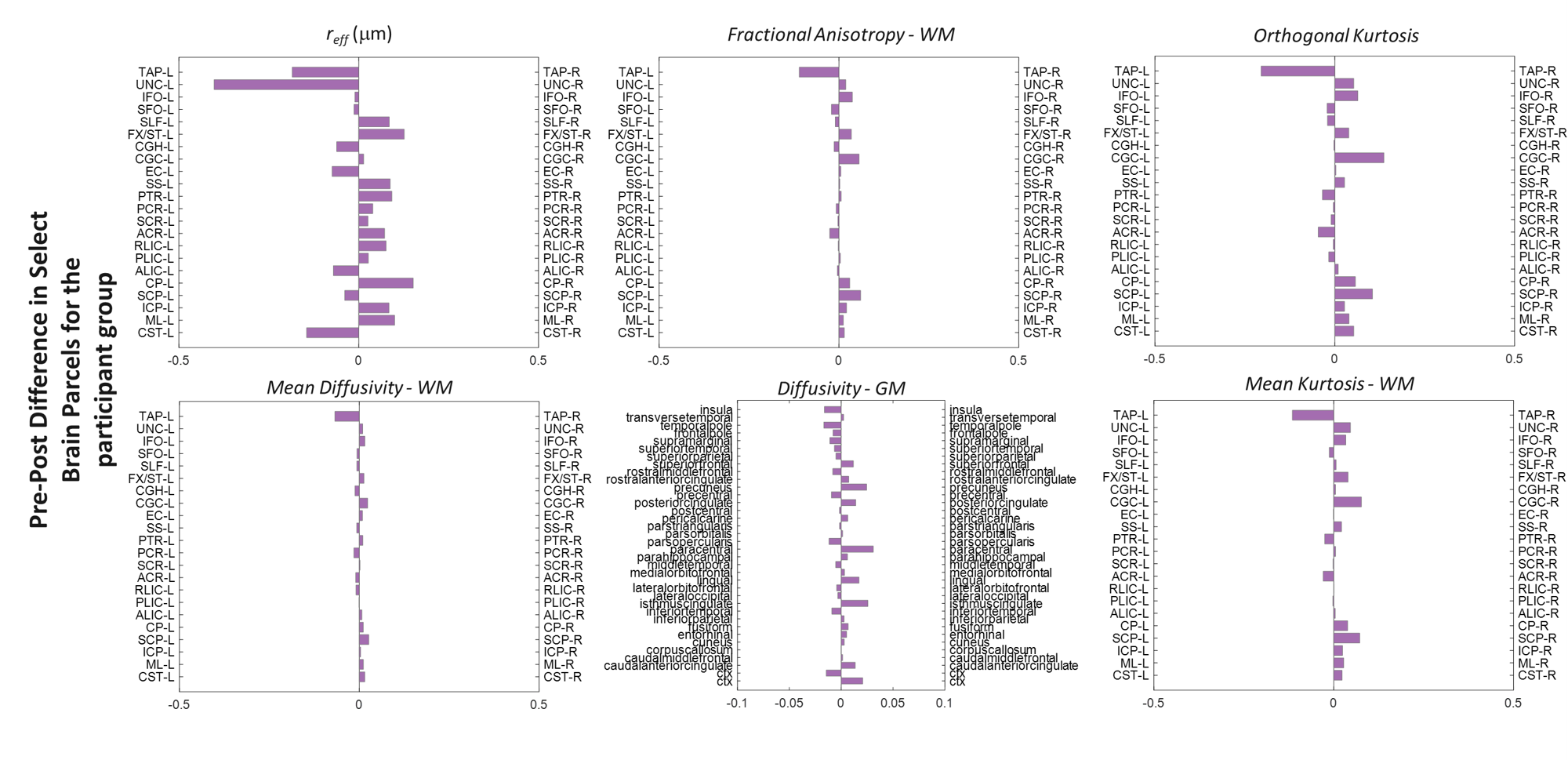
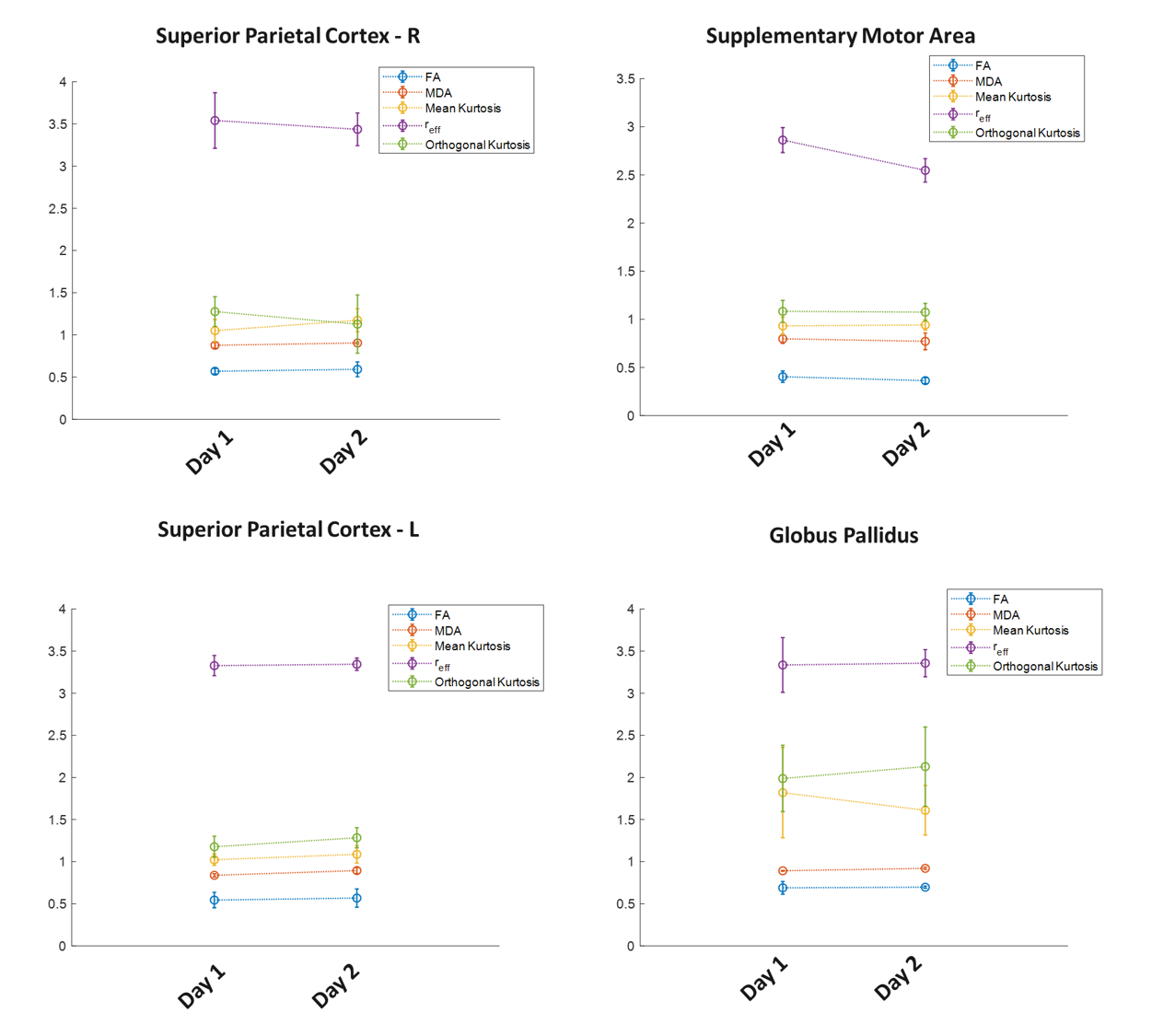
Analysis: A two-pronged approach was undertaken:1)WM and GM parcellation using JHU-181 and MNI-152 atlases; 2)Targeted ROIs based on prior literature[1].
Results & Discussion
Behavioral statistics are presented in Figure 3. With this pilot study, though participant variability was high, participants improved across blocks and, between days, as indicated by decreased lag (i.e., greater synchrony). However, likely due to the limited sample size and dominance of noise in group-statistics, there was no evidence for offline consolidation. Group differences (Figure 4) highlight regions of interest, the size and functional heterogeneity of large(r) parcels – in combination with the small heterogeneous sample – mask more specific neuroplastic effects.Targeted ROI’s from structures identified in prior group studies, are reported in Figure 5. Consistent with prior studies[1], the SMA highlights the largest difference in group-level metrics, with more localized changes in the globus pallidus and superior parietal cortex. Interestingly, reff exhibited the greatest plastic change in SMA. This is consistent with the observed decreasing trend in FA in the same region, and understandable given that this metric is also confounded by modifications to the extra-axonal space.
Conclusion
Preliminary results from the present limited pilot study reinforce the findings that rapid remodeling in response to cognitive experiences can be detected non-invasively with MRI. The higher sensitivity accorded by MAGNUS allowed for a limited sample size to elucidate microstructural changes compared to prior studies that have required larger sampling sizes(n≥20).Acknowledgements
CJS was supported by the Natural Sciences and Engineering Research Council (NSERC: RGPIN-2020-06812, DGECR-2020-00146) and the Heart and Stroke Foundation of Canada New Investigator Award and Catalyst from the Canadian Institutes of Health Research (HNC 170723).References
1. Tremblay, S.A., et al., White matter microstructural changes in short-term learning of a continuous visuomotor sequence. Brain Struct Funct, 2021. 226(6): p. 1677-1698.
2. Sagi, Y., et al., Learning in the fast lane: new insights into neuroplasticity. Neuron, 2012. 73(6): p. 1195-203.
3. Taubert, M., et al., Dynamic properties of human brain structure: learning-related changes in cortical areas and associated fiber connections. J Neurosci, 2010. 30(35): p. 11670-7.
4. Metzler-Baddeley, C., et al., Dynamics of White Matter Plasticity Underlying Working Memory Training: Multimodal Evidence from Diffusion MRI and Relaxometry. J Cogn Neurosci, 2017. 29(9): p. 1509-1520.
5. Caeyenberghs, K., et al., Dynamics of the Human Structural Connectome Underlying Working Memory Training. J Neurosci, 2016. 36(14): p. 4056-66.
6. Foo, T.K.F., et al., Highly efficient head-only magnetic field insert gradient coil for achieving simultaneous high gradient amplitude and slew rate at 3.0T (MAGNUS) for brain microstructure imaging. Magnetic Resonance in Medicine, 2020. 83(6): p. 2356-2369.
7. Weiger, M., et al., A high-performance gradient insert for rapid and short-T(2) imaging at full duty cycle. Magn Reson Med, 2018. 79(6): p. 3256-3266.
8. Huang, S.Y., et al., Connectome 2.0: Developing the next-generation ultra-high gradient strength human MRI scanner for bridging studies of the micro-, meso- and macro-connectome. Neuroimage, 2021. 243: p. 118530.
9. Kundu, P., et al., Differentiating BOLD and non-BOLD signals in fMRI time series using multi-echo EPI. Neuroimage, 2012. 60(3): p. 1759-70.
10. Newitt, D.C., et al., Gradient nonlinearity correction to improve apparent diffusion coefficient accuracy and standardization in the american college of radiology imaging network 6698 breast cancer trial. J Magn Reson Imaging, 2015. 42(4): p. 908-19.
11. Sperl, J.I., et al., Model-based denoising in diffusion-weighted imaging using g
eneralized spherical deconvolution. Magnetic Resonance in Medicine, 2017. 78(6): p. 2428-2438. 12. Doucette, J., C. Kames, and A. Rauscher, DECAES - DEcomposition and Component Analysis of Exponential Signals. Z Med Phys, 2020. 30(4): p. 271-278.
13. Sprenger, T., et al., Real valued diffusion-weighted imaging using decorrelated phase filtering. Magn Reson Med, 2017. 77(2): p. 559-570.
14. Veraart, J., et al., The variability of MR axon radii estimates in the human white matter. Human Brain Mapping, 2021. 42(7): p. 2201-2213.
Figures