2444
Diffusion-weighted GRASE sequence with 3D navigator for high-resolution time-dependency measurements in the human cortical gray matter1Key Laboratory for Biomedical Engineering of Ministry of Education, Department of Biomedical Engineering, College of Biomedical Engineering & Instrument Science, Zhejiang University, Hangzhou, China, 2Siemens Healthineers Ltd, Hangzhou, China
Synopsis
Keywords: Diffusion Acquisition, Brain
Motivation: Oscillating gradient sequences enable diffusion measurement at short diffusion-time (td), but it suffered from low resolution and SNR on clinical systems.
Goal(s): To explore the td-dependency in sophisticated structures, like cortical gray matter
Approach: We proposed a 3D navigator-based 3D gradient spin-echo (GRASE) sequence for whole-brain td–dMRI at 1.5 mm isotropic resolution, which enabled us to depict the cerebral cortex.
Results: We unveiled unique td-dependency patterns across cortical regions with different diffusion dispersion exponent (θ) based on the power-law, which was higher in the sensory cortex, like the occipital region, and lower in the high-order cortex like the temporal region.
Impact: The high-resolution td-dMRI technique provided a new approach to characterize the cortical micro-environment and is potentially useful to capture changes due to neurological diseases.
Introduction
Oscillating gradient (OG)1 stands as a pivotal method to evaluate time-dependent diffusion, particularly at short diffusion times (td), which promises insights into the intricate details of tissue microstructures2, 3. However, OG diffusion MRI (dMRI) suffered from low resolution and SNR on clinical systems, and thus, probing td-dependency in fine structures, like cortical gray matter (cGM), is difficult. 3D dMRI sequences4, 5 offered a solution to leverage high resolution and SNR. Nevertheless, whole-brain high-resolution 3D dMRI with multi-shots remains challenging due to the phase errors from eddy current and motion.Here, we developed a 3D multi-shot OGprep-GRASE equipped with a 3D navigator, to enable robust multi-shot whole-brain dMRI acquisitions. Moreover, we incorporated the GeneRalized Autocalibrating Partial Parallel Acquisition (GRAPPA)6 in both phase encoding directions for sequence acceleration. The td-dependency was meticulously quantified in the cGM within the human brain.
Methods
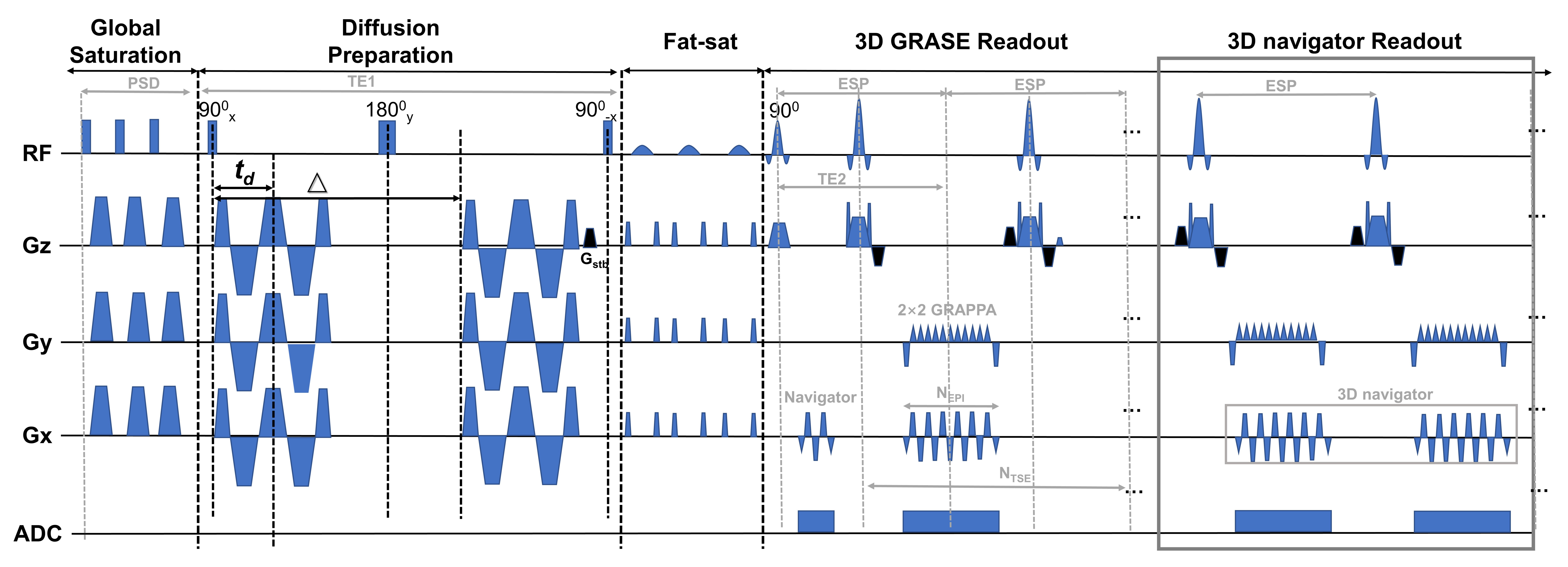
Pulse sequence: The 3D navigator-based 3D multi-shot OGprep-GRASE sequence was architecturally structured around five distinct modules: global saturation, diffusion preparation, fat saturation, 3D GRASE readout, and 3D navigator readout (Figure 1).Data acquisition: Seven healthy volunteers were enrolled with IRB approval. All scans were executed on a 3T Siemens Prisma scanner with a 64-channel head coil. The parameters were: TR/TE = 2600/136 ms, NEPI = 99, NTSE = 4 with 12shots, FOV = 220 × 200 mm2, 1.5 mm isotropic, GRAPPA accelerate factor = 2 × 2. Pulsed gradient (PG)-encodings at Δeff = 20ms, 30ms, 40ms, and OG-encodings at frequencies of 25 Hz and 50 Hz (effective td = 10ms and 5ms). b = 500 s/mm2 with 6 directions two averages and four b0.
Data analysis: Raw K-space datasets underwent GRAPPA-based reconstruction coupled with a 3D navigator-driven phase correction. We also acquired a 3D T1-weighted image and the cGM regions were segmented using it utilizing the AAL atlas7, which was co-registered to dMRI datasets. ADC measurements from multi-td data were obtained in each GM parcel to estimate the diffusion dispersion exponent (θ) based on the power-law model8: Dt =c×tdθ +Dt0. Higher θ indicated lower structural disorder according to the theory. To understand its microstructural basis, we compared θ with several other dMRI-based microstructural features from Neurite Orientation Dispersion and Density Imaging (NODDI) and Diffusion Kurtosis Imaging (DKI). We used 10 healthy subjects, from the HCP dataset, and calculated the averaged NODDI indices9 (encompassing the intra-cellular volume fraction (ICVF) and orientation dispersion index (ODI)) and DKI metrics10 (axial, mean, and radial kurtosis - AK, MK, and RK).
Results
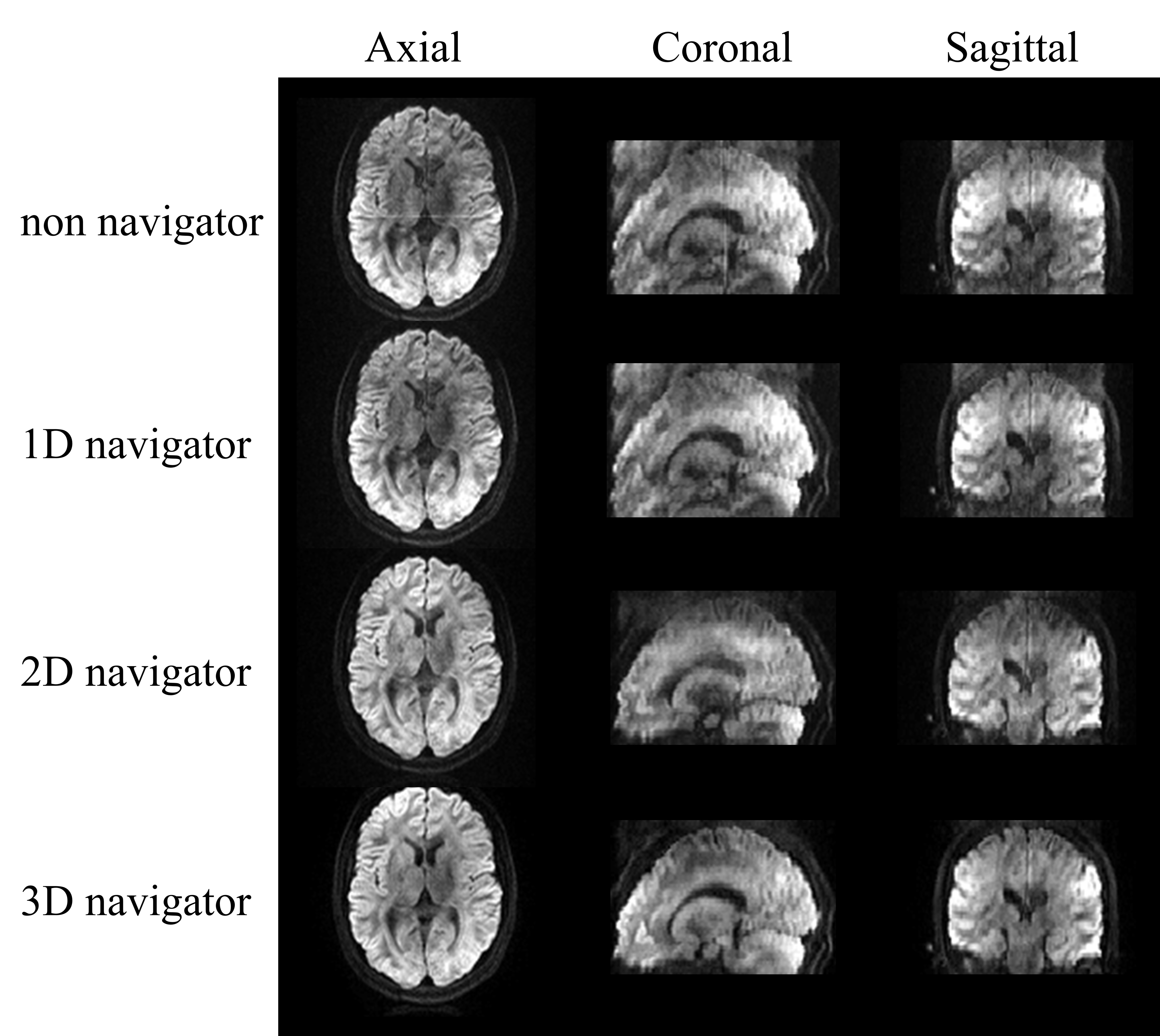
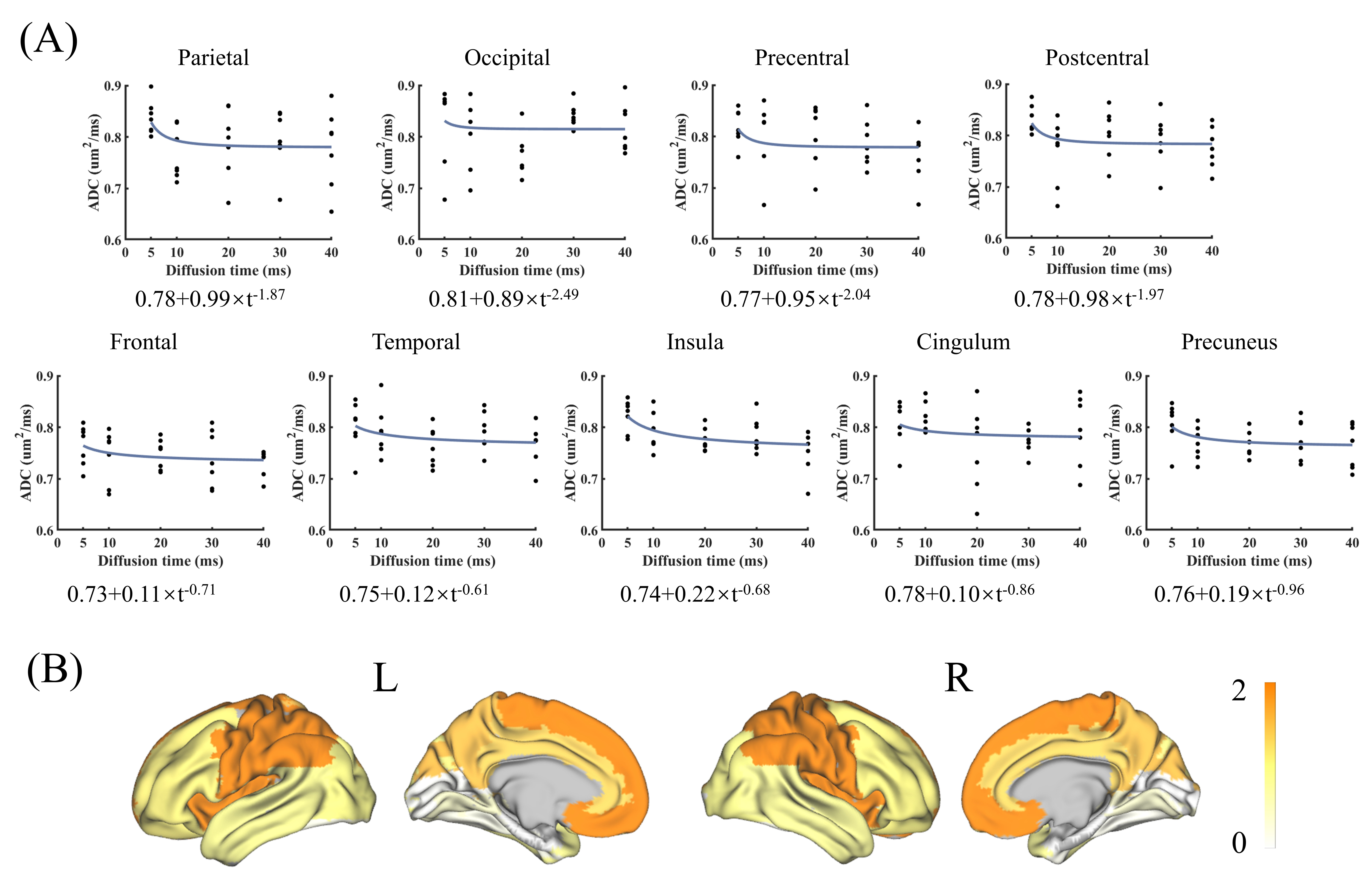
Figure 2 shows the DWIs obtained through the 3D navigator-based 3D-GRASE sequence at the b-value of 1000 s/mm2 in axial, coronal, and sagittal views, before and after 1D, 2D, and 3D navigator-based phase correction. Phase inconsistencies were substantially mitigated after navigator corrections.Using the 3D navigator-based sequence at 1.5 mm isotropic resolution, the td-dependent changes of ADC were quantified in several cGM regions (Figure 3(A)). Power-law analysis showed a θ exceeding 0.5 across all ROIs, which was the highest in the occipital lobe and lowest in the temporal lobes. Figure 3(B) showed the cGM mapping of θ, which further illustrated that the sensory cortex (e.g., parietal and occipital lobes) had the higher θ and the high-order cortex (e.g., frontal and temporal lobes) exhibited the lower θ, indicating higher structural disorder in the high-order cortex.
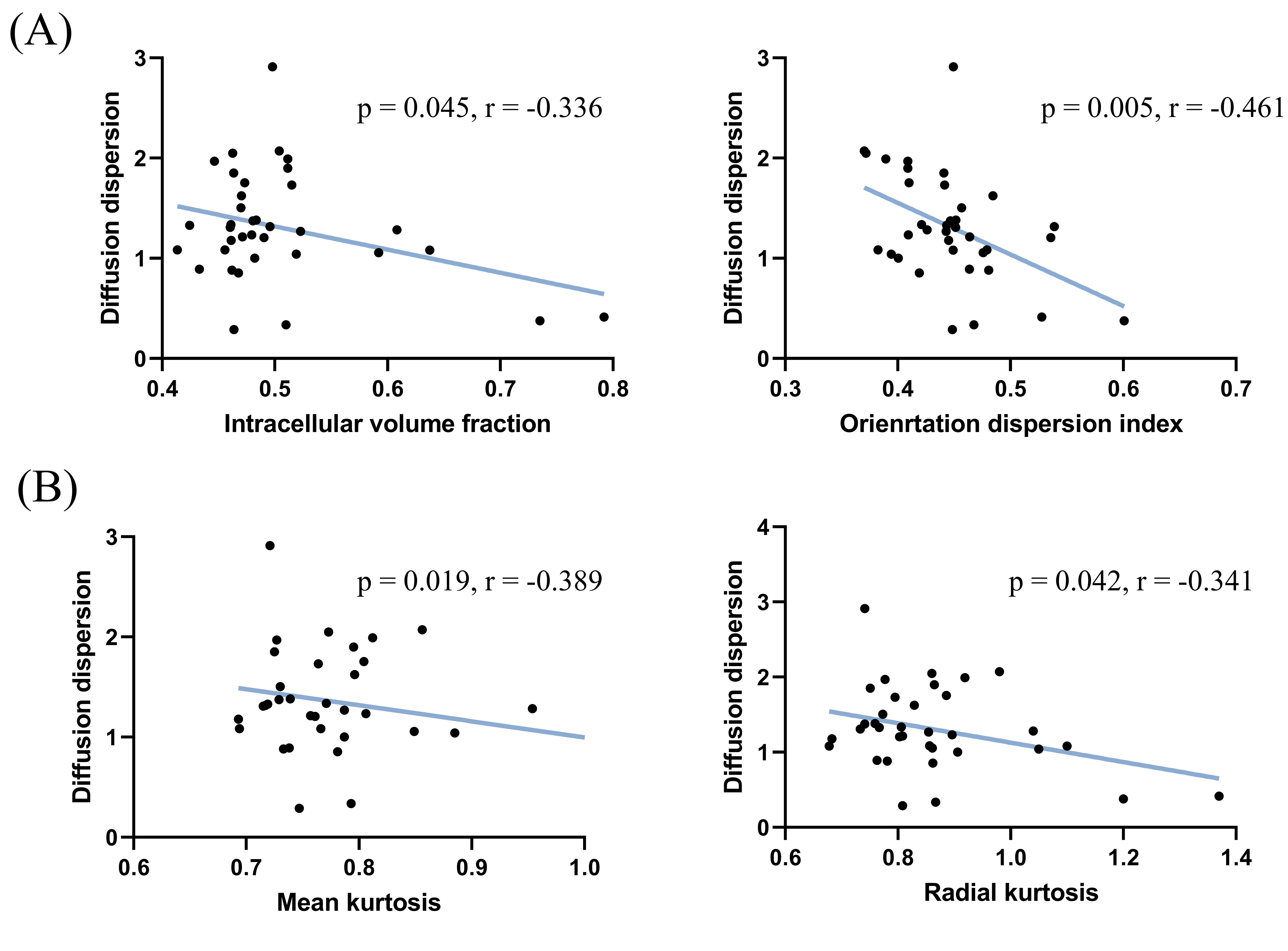
The Pearson correlation revealed negative associations between θ and NODDI indices (ICVF and ODI) (p<0.05, Figure 4(A)), suggesting higher neurite density and orientation disorder corresponded to higher structural disorder. Similar negative correlations were observed between θ and DKI indices (MK and RK) (Figure 4(B)), indicating higher non-Gaussian diffusion was related to higher structural disorder.
Discussion and Conclusion
This study introduced a 3D navigator-based 3D-GRASE sequence for high-resolution OG and PG-dMRI of the whole-brain on a clinical 3T system and charaterized the td-dependency in the cGM regions. The 3D navigator approach effectively corrected the inter-shot phase errors and considerably improved the td-dependency measurements.Distinct td-dependency profiles were identified in the human brain across different regions. Consistent with the recent report8, 11, the diffusion dispersion model indicated the estimated θ > 0.5 in GM regions. Moreover, the lower θ in the high-order cortex indicated higher structural disorder in these areas than primary cortex. This was further corroborated by the negative Pearson correlations between θ and both NODDI and DKI indices, underscoring a scenario where complicated microstructural organization corresponded to higher neurite density, orientation dispersion, and non-Gaussian diffusion. The high-resolution td-dMRI technique provided a new approach to characterize the cortical micro-environment and is potentially useful to capture changes due to neurological diseases.
Acknowledgements
This work is supported by the National Natural Science Foundation of China (81971606, 82122032), and Science and Technology Department of Zhejiang Province (2022C03057, 202006140)References
1. Baron CA, Beaulieu C. Oscillating gradient spin‐echo (OGSE) diffusion tensor imaging of the human brain. Magnetic resonance in medicine 2014;72(3):726-736.
2. Ianuş A, Jespersen SN, Duarte TS, et al. Accurate estimation of microscopic diffusion anisotropy and its time dependence in the mouse brain. Neuroimage 2018;183:934-949.
3. Arbabi A, Kai J, Khan AR, et al. Diffusion dispersion imaging: mapping oscillating gradient spin‐echo frequency dependence in the human brain. Magnetic resonance in medicine 2020;83(6):2197-2208.
4. Li H, Hsu Y-C, Zu T, et al. 3D diffusion MRI with Twin-navigator-based GRASE for comparison of fiber-tracking using 2D and 3D sequences in human whole brain.
5. Wu D, Liu D, Hsu YC, et al. Diffusion‐prepared 3D gradient spin‐echo sequence for improved oscillating gradient diffusion MRI. Magnetic resonance in medicine 2021;85(1):78-88.
6. Griswold MA, Jakob PM, Heidemann RM, et al. Generalized autocalibrating partially parallel acquisitions (GRAPPA). Magnetic Resonance in Medicine: An Official Journal of the International Society for Magnetic Resonance in Medicine 2002;47(6):1202-1210.
7. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002;15(1):273-289.
8. Lee H-H, Papaioannou A, Novikov DS, et al. In vivo observation and biophysical interpretation of time-dependent diffusion in human cortical gray matter. Neuroimage 2020;222:117054.
9. Zhang H, Schneider T, Wheeler-Kingshott CA, et al. NODDI: practical in vivo neurite orientation dispersion and density imaging of the human brain. Neuroimage 2012;61(4):1000-1016.
10. Veraart J, Sijbers J, Sunaert S, et al. Weighted linear least squares estimation of diffusion MRI parameters: strengths, limitations, and pitfalls. Neuroimage 2013;81:335-346.
11. Olesen JL, Østergaard L, Shemesh N, et al. Diffusion time dependence, power-law scaling, and exchange in gray matter. NeuroImage 2022;251:118976.
Figures