2410
Voxel size matter: an analysis of the sampling bias in Monte Carlo simulation and realistic synthetic substrates using CACTUS1LTS5, Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Switzerland, 2Radiology, Centre Hospitalier Universitaire Vaudois and University of Lausanne, Lausanne, Switzerland, 3CIBM Center for Biomedical Imaging, Lausanne, Switzerland, 4Computer Science, Université de Sherbrooke, Sherbrooke, QC, Canada
Synopsis
Keywords: Diffusion Modeling, Simulations, White-Matter, Monte-Carlo simulations.
Motivation: This work addresses the sampling bias related to voxel size and boundary conditions in DW-MRI Monte-Carlo simulations.
Goal(s): The study aims to understand how voxel-size and boundary-conditions influence Monte-Carlo simulations in DW-MRI to ensure convergence and minimise errors.
Approach: It uses simulations with realistic synthetic white-matter substrates and calculates diffusion propagators and apparent diffusion coefficients to measure simulation accuracy.
Results: It underscores the significance of voxel-size and boundary conditions in Monte-Carlo simulation and offers insights for better simulation parameters in DW-MRI. It also highlights conditions where errors can reach 20% and shows the need for larger voxel-sizes to achieve convergence.
Impact: The analysis focuses on making Monte-Carlo simulations reliable, enabling model validation from DW-MRI techniques. This has the potential to substantially improve microstructure assessment precision in clinical practice, enabling more accurate DW-MRI analysis.
Introduction
Monte-Carlo(MC) simulations are invaluable for analysing microstructure models9 of Diffusion-Weighted Magnetic Resonance Imaging(DW-MRI). They emulate the Brownian motion dynamics of water molecules within a three-dimensional mesh representing virtual tissue geometries and characterise the diffusion process in complex substrates without analytical solutions. However, a key challenge is ensuring the accuracy of DW-MRI signals from Monte-Carlo1,8 simulations: different boundary conditions and voxel sizes on the same substrate could introduce non-negligible signal variations. This abstract explores the impact of voxel size and boundary conditions on simulation accuracy. We analyse the sampling bias inherent in Monte-Carlo simulations when not properly curated. In addition, we provide guidelines for choosing the adequate voxel size to ensure the diffusion propagator’s convergence.Methods
Monte-Carlo simulations and diffusion propagators in virtual substrates: We generated two synthetic white-matter virtual-substrates using CACTUS1,12 with voxel-size=220x220x220µm³ and intracellular volume fractions(ICVF) of 65% and 85%. These substrates comprised synthetic fibres with a mean radius11 of 0.6 µm, 0.7 average g-ratio14, 5° global dispersion10, and fiber-radii drawn from a gamma-distribution11 Γ(κ=5.05,θ=0.13).The diffusion propagators were obtained using particle trajectories from the MC-DC simulator1. We initiated particles in intra- and extra-fiber spaces, excluding those between inner and outer diameters to exclude the myelin-water-compartment2,3. We generated two propagators for each substrate with diffusivities of D=0.6x10⁻³mm²/s (ex-vivo18) and D=3.0x10⁻³mm²/s (in-vivo16,17), with the same D assumed for intra- and extra-cellular compartments. All particles underwent 10,000 diffusion steps with diffusion time Δ=58ms1,2,5,6. Each experiment was repeated ten times to assess variability. In the simulations, we maintained a particle density of 1 particle per µm³, following1,3,4. We also created a reference gold standard with a higher seeding density of 20 particles per µm³.
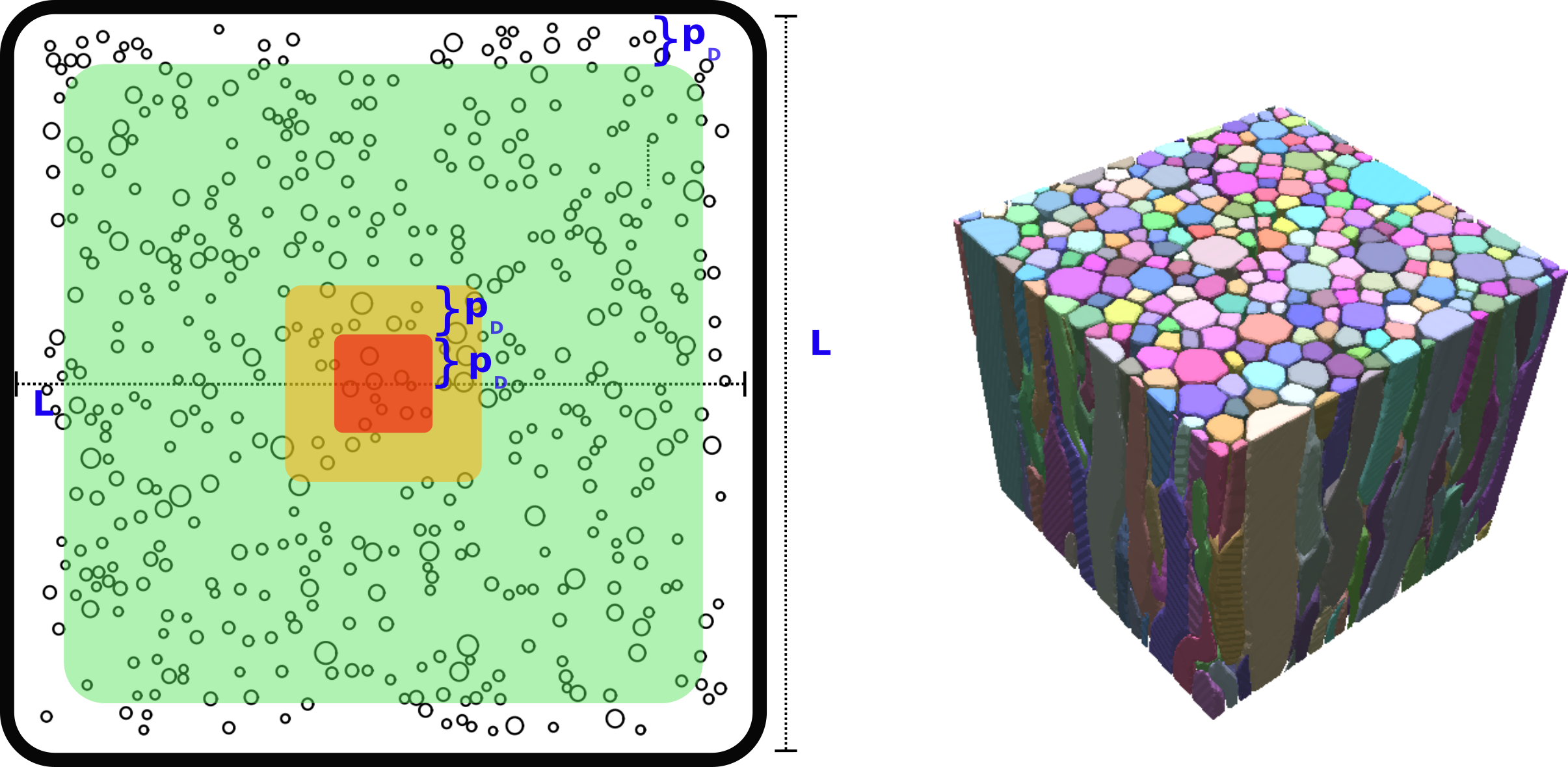
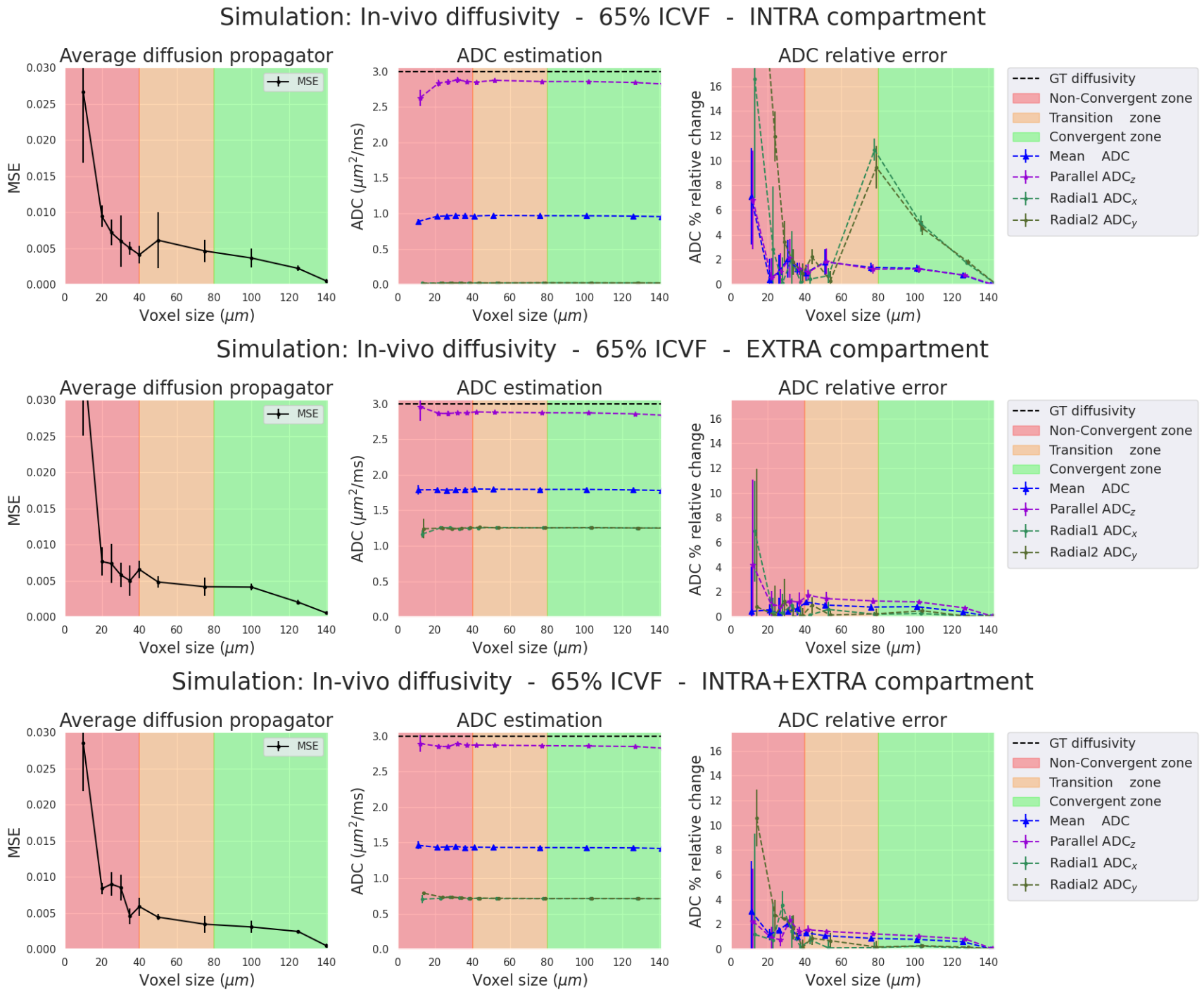
Voxel sizes and zones of interest: To prevent particles from having boundary problems, we used smaller volumes of (Lmax)3 (Lmax=180µm for ex-vivo and 140µm for in-vivo experiments). These Lmax-sized voxels served as the largest voxel size for the subsequent simulations. We computed particle trajectories inside ‘virtual’ voxels ranging from L=10µm to Lmax. For interpretation, we categorised the biggest voxels into three zones of interest based on their distance from the centre of the voxel (Figure-1): non-convergent [0,pD], transition [pD,2pD], and convergent [2pD,Lmax), with pD denoting the distance at which 95% of particles in a free medium travel a distance smaller than pD during the diffusion time. In ex-vivo pD ~ 20µm and in-vivo pD ~ 40µm.
Evaluation: We create the average diffusion propagator using 500 sphere directions, each representing mean squared displacement in its respective direction. We quantify differences between the gold standard and reduced voxel average diffusion propagators using mean-square error (MSE). Moreover, we computed the relative errors of the apparent-diffusion-coefficient(ADC) derived from the propagators, including 1) the mean ADC, 2) Parallel ADC along the main fibres axis, and 3) Radial ADCs along the x- and y-axes perpendicular to the fibres. ADCs were calculated using Einstein's equation7 <(X(Δ)-X(0))2>=(2nADCΔ), with n=3 for mean ADC and n=1 for parallel and radial ADCs.
Results and Discussion
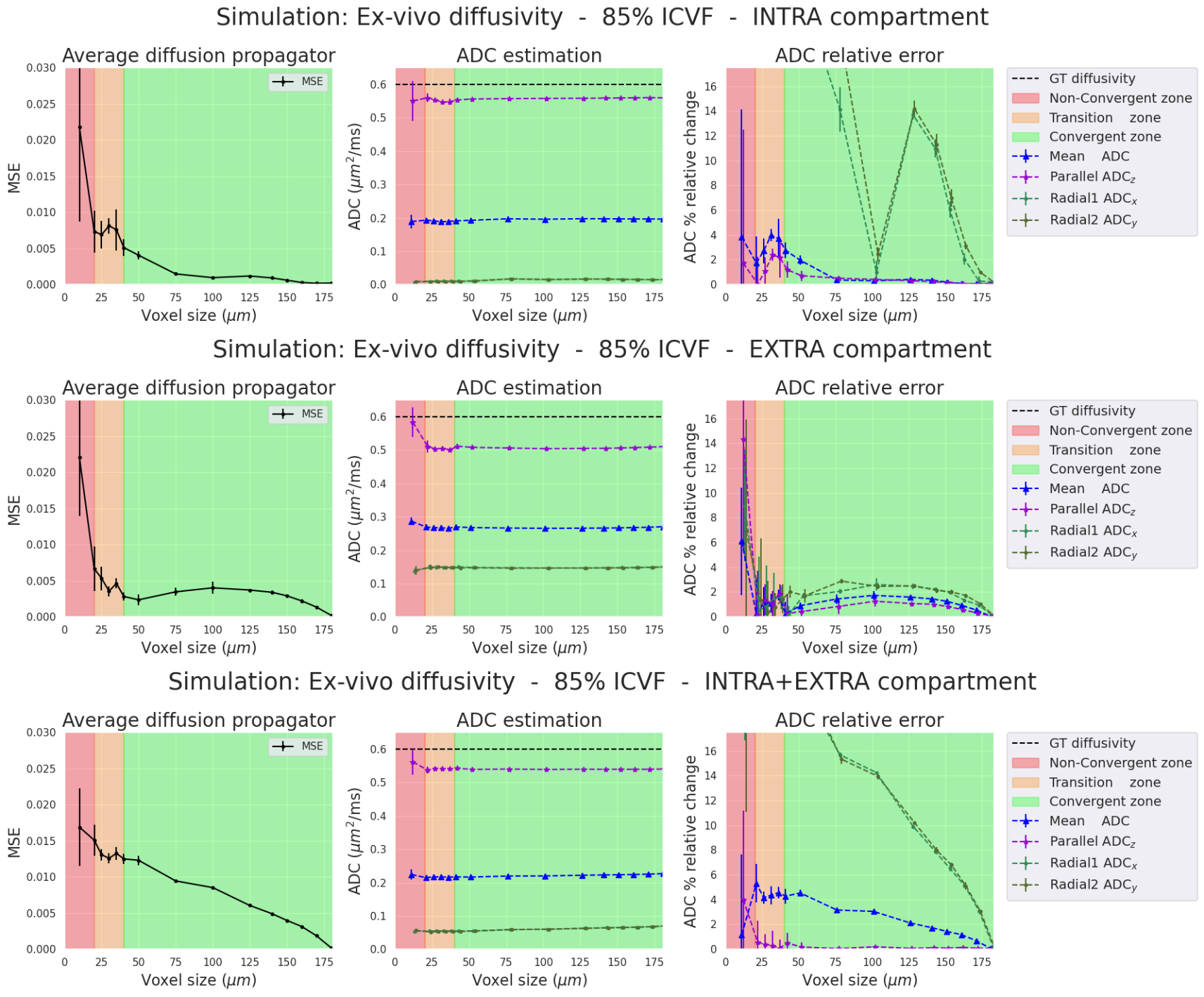
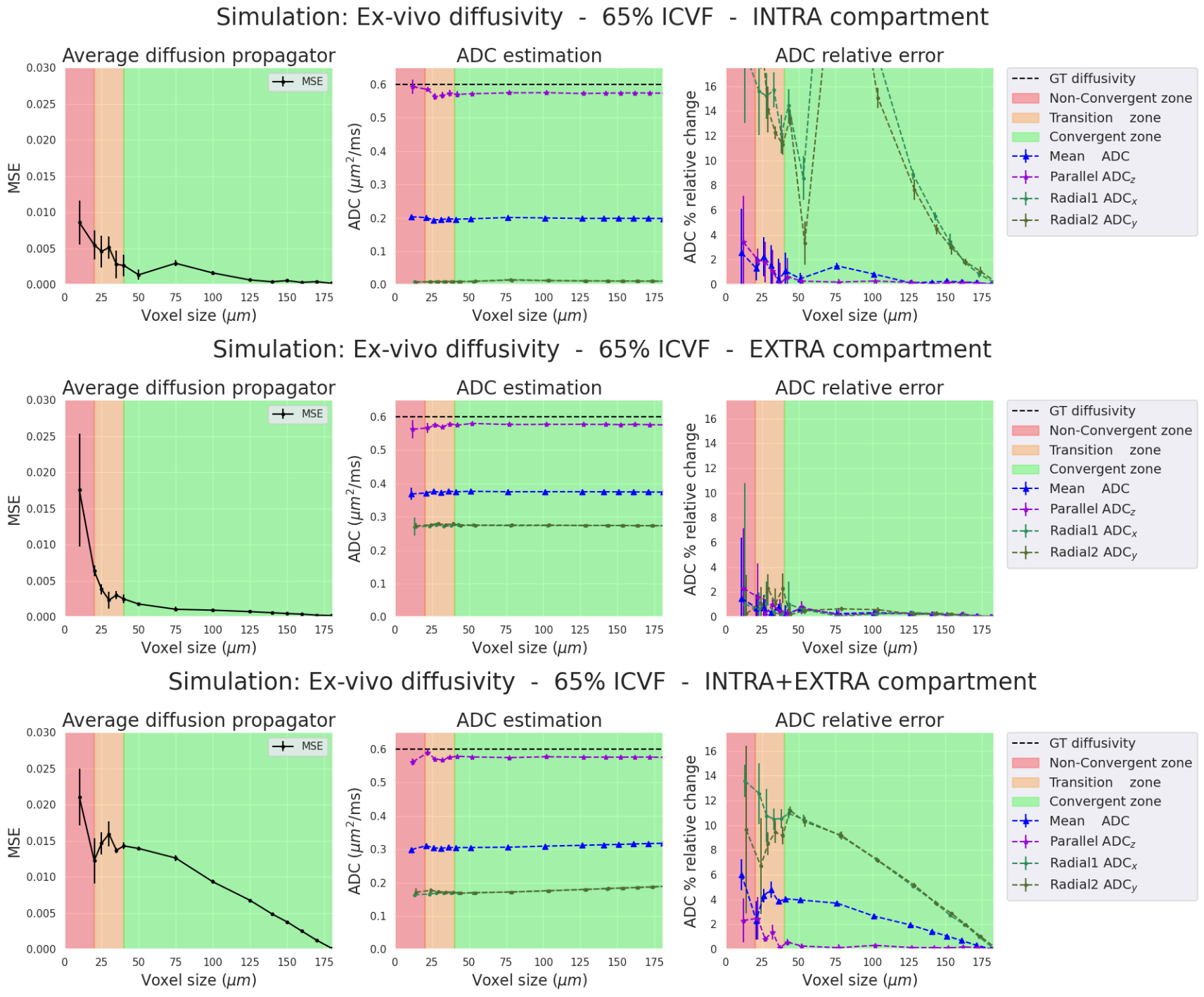
The propagator convergence, the ADC and relative errors are presented for the four simulations (two diffusivities and two ICVF). For each simulation, we separate the analysis into three cases: 1) intra-cellular(IC) particles, 2) extra-cellular(EC) particles, and 3) both compartments(IC+EC).Ex-vivo diffusivity simulations (Figures 2-3). In all three cases, the ADC relative errors exceed 30% for voxel size within the "Non-Convergent” and “Transition” zones. The error was reduced below 3% inside the "convergent” zone. Also, the estimated parallel and mean ADCs closely matched the ground truth values for the intra- and extra-cellular cases (2.5% relative error in the “convergent” zone”). In the intra-cellular case, the radial ADC estimation was small (0.025 mm²/s) due to the small axon sizes, showing a relative error of 30%. As expected, high relative errors could arise when estimates approach zero.
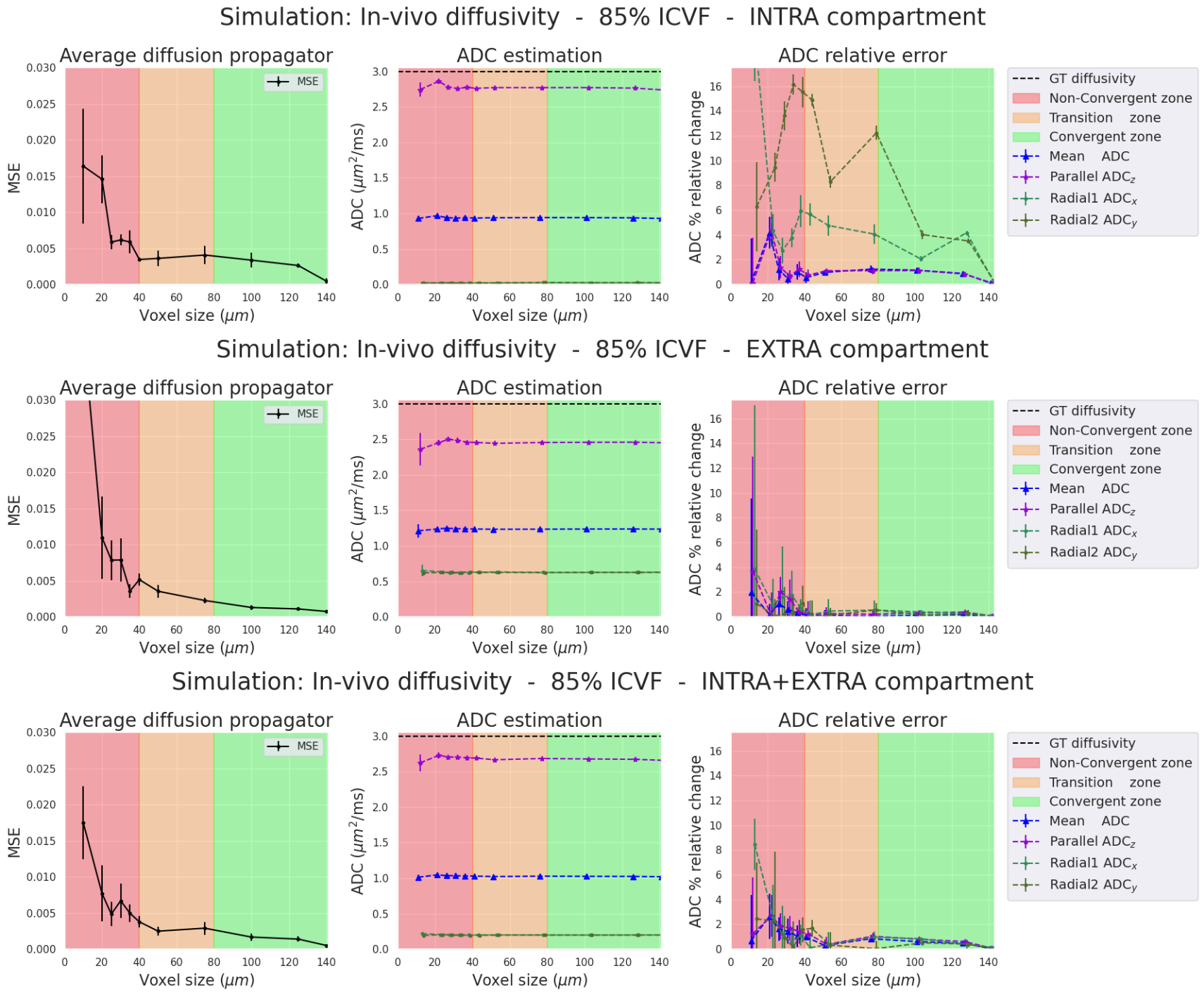
In-vivo diffusivity Simulations (Figures 4-5). Results between in-vivo and ex-vivo cases are comparable in their respective zones, which are functions of the pD, meaning that bigger voxels are needed for higher diffusivities. In summary, our experiments show that we require voxel sizes larger than 2*pD to ensure the stability and convergence of Monte-Carlo simulations, both ex-vivo and in-vivo.
Conclusion
Our study explores the impact of voxel size and boundary conditions on simulation accuracy. The diffusivities in the simulations affect the propagator convergence and introduce bias into apparent diffusion estimates. Our results highlight the importance of an adequate voxel size (bigger than 2*pD) given the employed simulation parameters (simulation time and diffusivity), to minimise the propagator’s estimation error and ensure accurate convergence. Further research is essential to explore DW-MRI signal sensitivity with respect to the propagator sensibility.Acknowledgements
This work is partly supported by the Swiss National Science Foundation (SNSF) under grant Nbr 205320_175974. We acknowledge access to the facilities and expertise of the CIBM Center for Biomedical Imaging, a Swiss research centre of excellence founded and supported by Lausanne University Hospital (CHUV), University of Lausanne (UNIL), Ecole Polytechnique Fédérale de Lausanne (EPFL), University of Geneva (UNIGE), Geneva University Hospitals (HUG), and Université de Sherbrooke. Erick J. Canales-Rodríguez was supported by the SNSF, Ambizione grant PZ00P2_185814.References
[1] Rafael-Patino, Jonathan, et al. "Robust Monte-Carlo simulations in diffusion-MRI: Effect of the substrate complexity and parameter choice on the reproducibility of results." Frontiers in neuroinformatics 14 (2020): 8.
[2] Villarreal-Haro JL, Gardier R, Canales-Rodríguez EJ, et al. CACTUS: a computational framework for generating realistic white matter microstructure substrates. Front Neuroinform. 2023;17:1208073.
[3] Rafael-Patino, J., Girard, G., Truffet, R., Pizzolato, M., Caruyer, E., & Thiran, J. P. (2021). The diffusion-simulated connectivity (DiSCo) dataset. Data in Brief, 38, 107429.
[4]Girard, G., Rafael-Patiño, J., Truffet, R., Aydogan, D. B., Adluru, N., Nair, V. A., ... & Thiran, J. P. (2023). Tractography passes the test: Results from the diffusion-simulated connectivity (disco) challenge. NeuroImage, 277, 120231.
[5] Hall, M. G., and Alexander, D. C. (2009). Convergence and parameter choice for monte-carlo simulations of diffusion MRI. IEEE Trans. Med. Imaging 28, 1354–1364. doi: 10.1109/TMI.2009.2015756
[6] Yeh, C.-H., Schmitt, B., Bihan, D. L., Li-Schlittgen, J.-R., Lin, C.-P., and Poupon, C. (2013). Diffusion microscopist simulator: a general monte carlo simulation system for diffusion magnetic resonance imaging. PLoS ONE 8:e76626. doi: 10.1371/journal.pone.0076626
[7] Einstein, A. (1956). Investigations on the Theory of the Brownian Movement. Courier Corporation.
[8] Lee, Hong-Hsi, Els Fieremans, and Dmitry S. Novikov. "Realistic Microstructure Simulator (RMS): Monte Carlo simulations of diffusion in three-dimensional cell segmentations of microscopy images." Journal of Neuroscience Methods 350 (2021): 109018.
[9] Novikov, Dmitry S., et al. "Quantifying brain microstructure with diffusion MRI: Theory and parameter estimation." NMR in Biomedicine 32.4 (2019): e3998.
[10] Andersson, M., Kjer, H. M., Rafael-Patino, J., Pacureanu, A., Pakkenberg, B., Thiran, J. P., ... & Dyrby, T. B. (2020). Axon morphology is modulated by the local environment and impacts the noninvasive investigation of its structure–function relationship. Proceedings of the National Academy of Sciences, 117(52), 33649-33659.
[11] Liewald, Daniel, et al. "Distribution of axon diameters in cortical white matter: an electron-microscopic study on three human brains and a macaque." Biological cybernetics 108.5 (2014): 541-557.
[12] Haro, Juan Luis Villarreal, et al. "Towards a high-density packing white matter substrate generator for Monte-Carlo simulations."
[13] Zhang, Hui, et al. "NODDI: practical in vivo neurite orientation dispersion and density imaging of the human brain." Neuroimage 61.4 (2012): 1000-1016.
[14] Stikov, Nikola, et al. "In vivo histology of the myelin g-ratio with magnetic resonance imaging." Neuroimage 118 (2015): 397-405.]
[15] Abdollahzadeh, Ali, et al. "DeepACSON automated segmentation of white matter in 3D electron microscopy." Communications biology 4.1 (2021): 1-14.
[16] Mills, R. (1973). Self-diffusion in normal and heavy water in the range 1-45. deg. The Journal of Physical Chemistry, 77(5), 685-688.
[17] Ledo, A., Fernandes, E., Salvador, A., Laranjinha, J., & Barbosa, R. M. (2022). In vivo hydrogen peroxide diffusivity in brain tissue supports volume signaling activity. Redox Biology, 50, 102250.
[18] Roebroeck, A., Miller, K. L., & Aggarwal, M. (2019). Ex vivo diffusion MRI of the human brain: Technical challenges and recent advances. NMR in Biomedicine, 32(4), e3941.
Figures