2406
Flexible and computationally efficient framework for diffusion MRI simulations in realistic neuron morphologies1Université de Lausanne, Lausanne, Switzerland, 2Lausanne University Hospital (CHUV), Lausanne, Switzerland, 3Cardiff University, Cardiff, United Kingdom, 4Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Switzerland
Synopsis
Keywords: Simulation/Validation, Gray Matter, Software tools, Diffusion software, Monte Carlo
Motivation: Analytical diffusion models are limited in the complexity of brain tissue they capture, and creating complex numerical phantoms using meshes can be arduous.
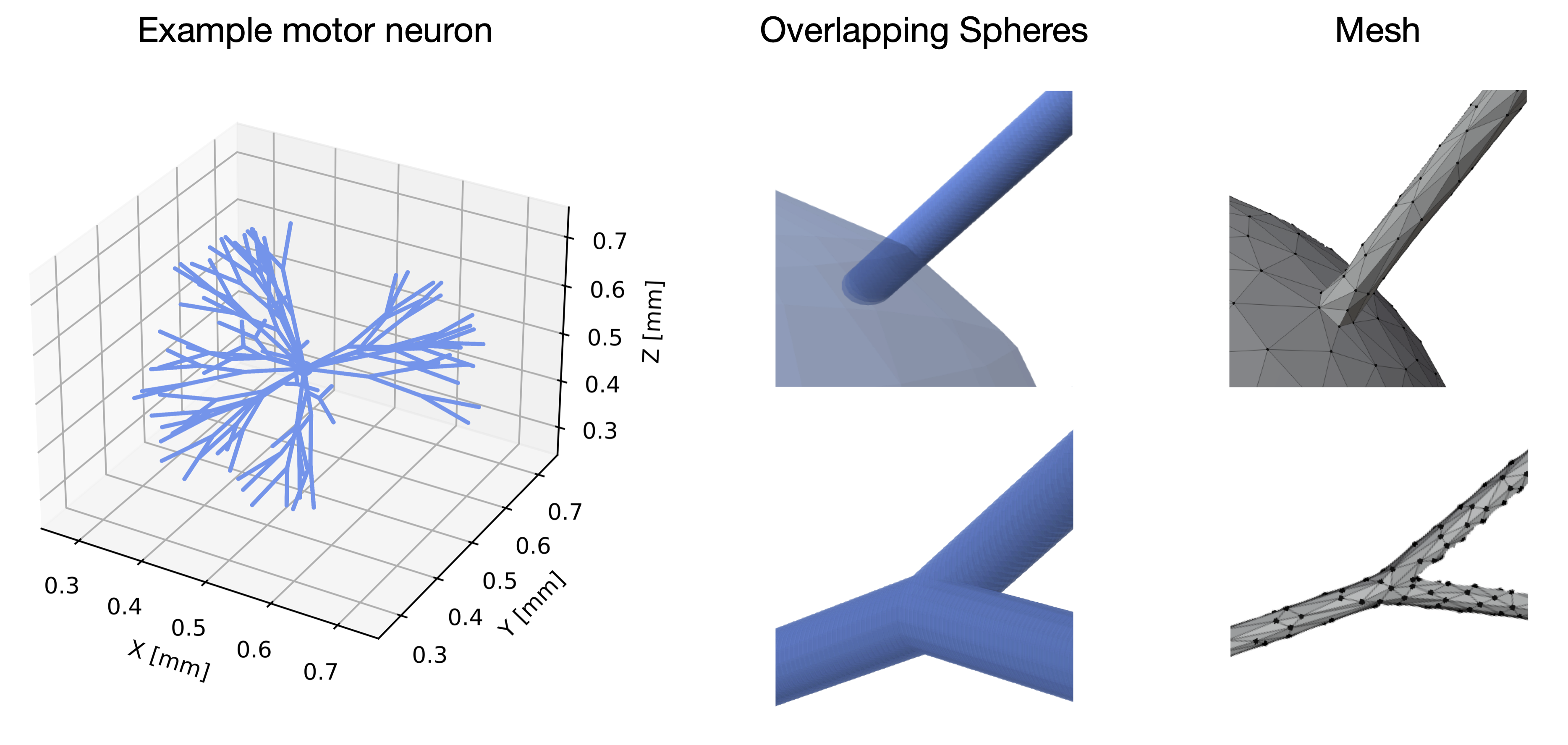
Goal(s): This work introduces an efficient and user-friendly software for generating realistic neurons using overlapping spheres.
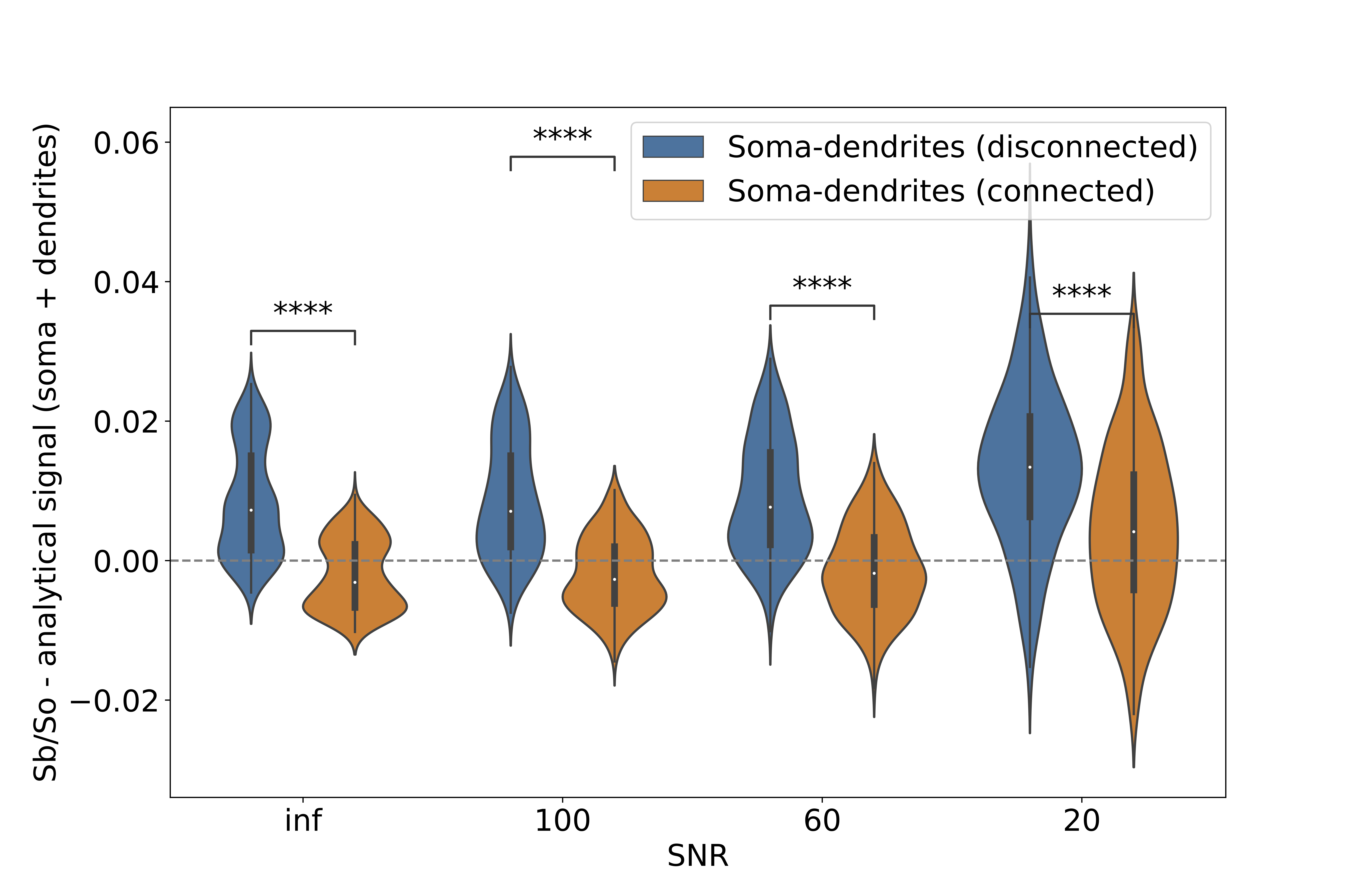
Approach: Through Monte-Carlo simulations, we evaluate the impact of sphere overlap, soma-dendrites water exchange, and SNR on dMRI signals.
Results: The findings indicate that modest sphere overlap maintains signal quality. Additionally, the soma-dendrites water exchange has a significant impact on the signal, even at realistic SNR. This work has promising implications for non-invasively quantifying gray matter microstructure, especially neuronal packing and cell membrane permeability.
Impact: Numerical phantoms of gray matter built with overlapping spheres offer an efficient and flexible way of studying water diffusion. This approach reduces the heavy meshing and cleaning process to trivial building blocks that can be adapted into realistic neuronal substrates.
Introduction
Given the limitation of analytical diffusion models to render the complexity of brain tissue, numerical phantom have been used for the flexibility to tune their properties to any level of realism1. Most phantoms mimic white matter2 using triangular meshes. This involves a substantial computational expense for the substrate generation. Building a regular manifold mesh that preserves the target proportions can be arduous. We propose an in-house easy-to-use software that generates realistic neurons using overlapping spheres at a reduced computational cost, as initially introduced in3. Using Monte-Carlo simulations (MCS), we investigate the impact on the dMRI signal of the overlap between adjacent spheres, soma-dendrites water exchange, and SNR. We further compare the simulation computation time between overlapping spheres vs mesh approach for one neuron.Methods
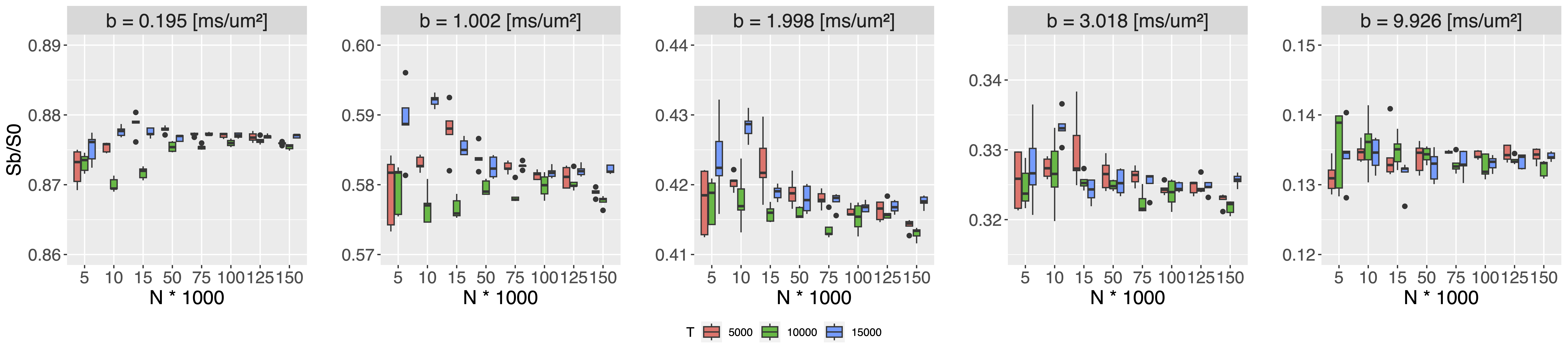
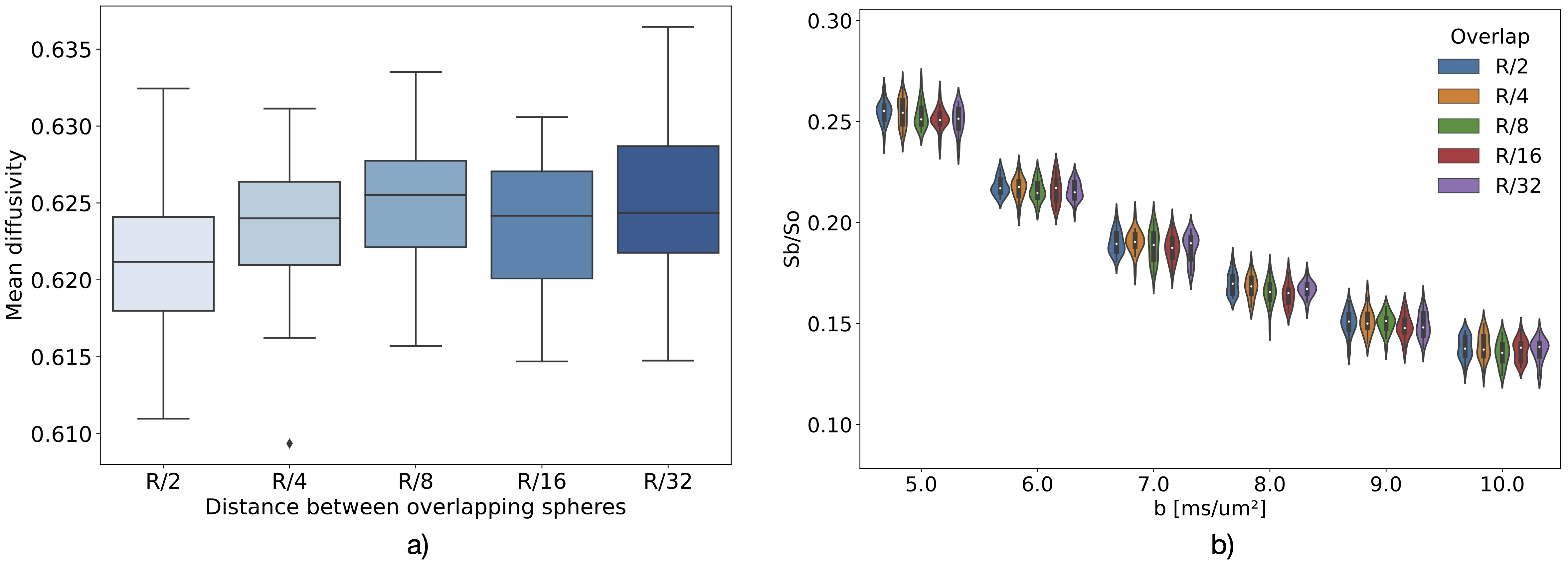
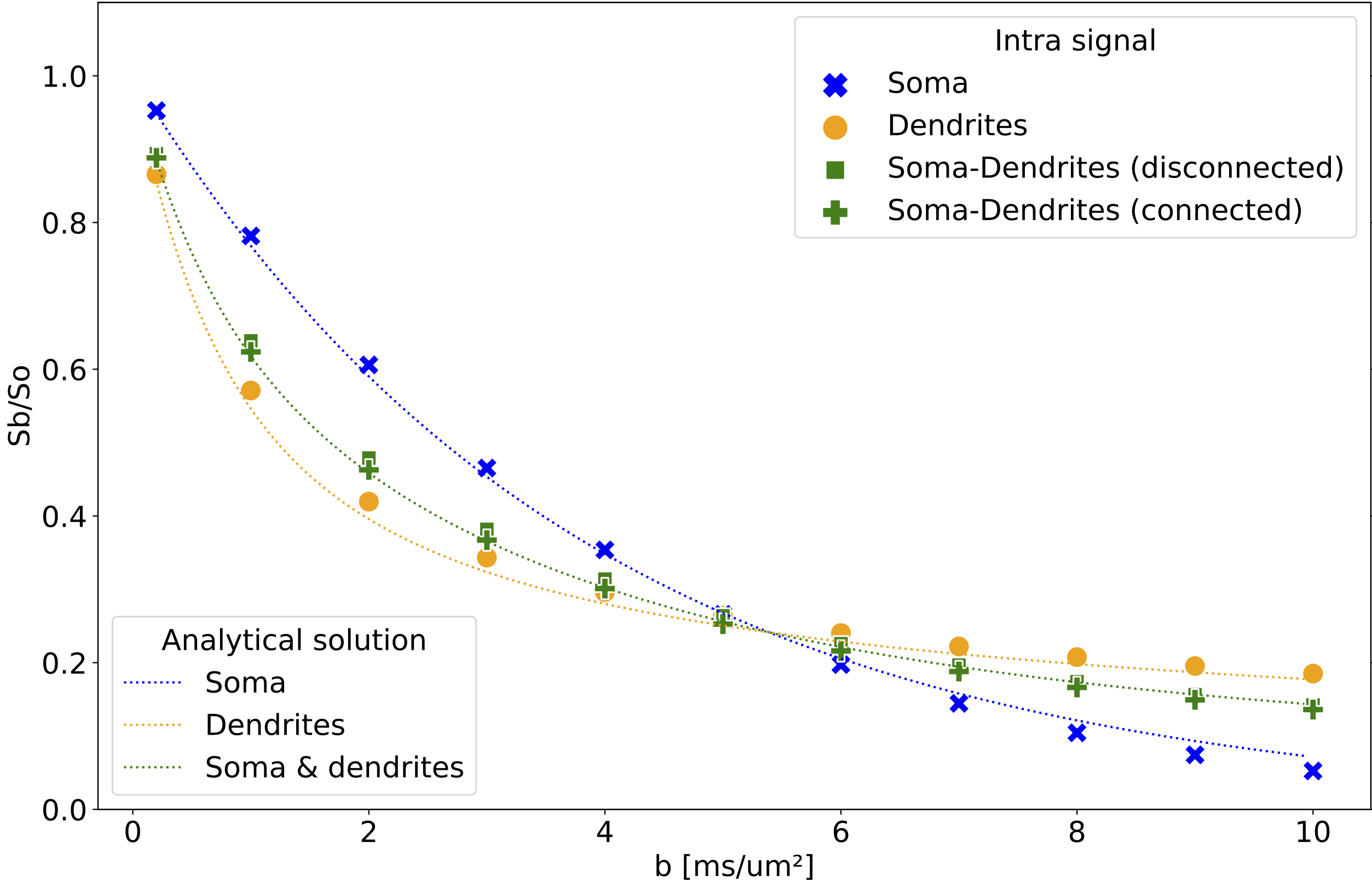
Neuron substrates were built using in-house software in C++ (Fig. 1) and MCS performed using a modified MCDC simulator4 to accommodate overlapping spheres. The convergence of the simulator propagators was benchmarked for different timesteps T and walkers N on an example motor neuron5 (Fig. 1). Each (T,N) combination was evaluated over 5 repetitions for a Pulse-Gradient Spin Echo (PGSE) sequence with Δ=50ms, δ=16.5ms, TE=67ms, b=[0, 0.2, 1:10] ms/μm², and 21 directions. Using the optimal (T,N) combination for signal convergence, we simulated diffusion within 5 neurons with a distance between spheres of R/2, R/4, R/8, R/16 and R/32. We probed diffusion in four environments (soma alone, dendrites alone, soma disconnected from dendrites, as modeled in6, and soma connected to dendrites) using the minimum overlap beyond which the signal converged. We compared the numerical powder-averaged dMRI signal to the analytical solutions for diffusion in a sphere7, in randomly-oriented infinite sticks8 (ROS), and a linear combination of the two. To study the connected and disconnected case at different SNR, Gaussian noise with σ=1/SNR (SNR=inf, 100, 60, 20) was added on the real and imaginary part of the signal. All computations were performed on a HPC using 1 node (48 CPUs, 1TB, 24 Cores, 1585.641MHz). The mesh was generated with BLENDER software using the “SWC Mesh” add-on that transforms SWC to mesh. A decimation factor of 5% was applied for mesh simplification.Results
Convergence was ensured for step length=0.259μm, N=50000 walkers (Fig. 2) and an overlap of R/4 (Fig.3). The computation time for an overlap of R/4 (89618 spheres) was 4.31±0.08min vs 2.86±0.04 min for a mesh with 48979 vertex and 97954 faces. The soma signal agrees with the analytical solution in the Gaussian phase approximation (GPA) up to b=6ms/μm², beyond which the numerical signal is lower, as expected from the analytical negative kurtosis9, (Fig. 4). The dendrites signal deviates from ROS at b < 5ms/μm² possibly due to finite segment length and branching, and the disconnected soma+dendrites follows the linear combination of analytical sphere+ROS. No matter the SNR, the signal was significantly lower in the connected vs disconnected soma+dendrites case (Figs.4-5) due to reduced restriction. For connected soma and dendrites, the final molecule density displayed an imbalance with 0.24% excess molecules in the dendrites vs soma (data not shown).Discussion
Based on the substrates and parameters used, an overlap of R/4 was sufficient to ensure signal convergence. This optimal overlapping factor should be reevaluated if using different diffusion time or characteristic lengths. Exchange between soma and dendrites has so far been neglected in models8 though it was shown to have an effect in straight but not in branching neurons10. Our simulations show this exchange also has an impact in branched neurons, even with the addition of realistic noise, but may become marginal vs exchange with an extracellular space. Remarkably, calculation times for the same neuron materialized using overlapping spheres vs mesh were comparable, although MCDC was optimized for meshes. Optimizing the simulator for overlapping spheres will potentially further reduce the computation time. Our framework has the advantage of eliminating the meshing step, for which leakage control and complexity optimization can be time-consuming. It also allows accurate dimensioning (vs substrates that shrink in the meshing process), and enables a straightforward deformation of the neuron, e.g. to model beading, swelling, etc.Conclusion
Diffusion simulations within neuronal substrates are a promising tool to non-invasively quantify GM microstructure in vivo. We use a flexible and computationally efficient framework to generate realistic neurons, where we show that intracellular water transit without any osmotic gradient between soma and dendrites has a detectable influence on the diffusion signal. Further developments will consist in packing neurons to yield a realistic extracellular space, considering dendrite tortuosity (facilitated by the sphere building blocks) and membrane permeability11.Acknowledgements
This work was supported by the ERC Starting Grant 'FIREPATH', SERI no. MB22.00032.References
1I. O. Jelescu, M. Palombo, F. Bagnato, and K. G. Schilling, “Challenges for biophysical modeling of microstructure,” Journal of Neuroscience Methods, vol. 344, p. 108861, Oct. 2020.
2E. Fieremans and H.-H. Lee, “Physical and numerical phantoms for the validation of brain microstructural MRI: A cookbook,” NeuroImage, vol. 182, pp. 39–61, Nov. 2018.
3K. Ginsburger, F. Matuschke, F. Poupon, J.-F. Mangin, M. Axer, and C. Poupon, “MEDUSA: A GPU-based tool to create realistic phantoms of the brain microstructure using tiny spheres,” NeuroImage, vol. 193, pp. 10–24, June 2019.
4J. Rafael-Patino, D. Romascano, A. Ramirez-Manzanares, E. J. Canales-Rodríguez, G. Girard, and J.-P. Thiran, “Robust Monte-Carlo Simulations in Diffusion-MRI: Effect of the Substrate Complexity and Parameter Choice on the Reproducibility of Results,” Frontiers in Neuroinformatics, vol. 14, 2020.
5M. Palombo, D. C. Alexander, and H. Zhang, “A generative model of realistic brain cells with application to numerical simulation of the diffusion-weighted MR signal,” NeuroImage, vol. 188, pp. 391–402, Mar. 2019.
6M. Palombo, A. Ianus, M. Guerreri, D. Nunes, D. C. Alexander, N. Shemesh, and H. Zhang, “SANDI: A compartment-based model for non-invasive apparent soma and neurite imaging by diffusion MRI,” NeuroImage, vol. 215, p. 116835, July 2020.
7J. S. Murday and R. M. Cotts, “Self-Diffusion Coefficient of Liquid Lithium,” The Journal of Chemical Physics, vol. 48, pp. 4938–4945, Sept. 2003.
8P. T. Callaghan, K. W. Jolley, and J. Lelievre, “Diffusion of water in the endosperm tissue of wheat grains as studied by pulsed field gradient nuclear magnetic resonance.,” Biophysical Journal, vol. 28, pp. 133–141, Oct. 1979.
9H.-H. Lee, E. Fieremans, and D. S. Novikov, “What dominates the time dependence of diffusion transverse to axons: Intra- or extra-axonal water?,” NeuroImage, vol. 182, pp. 500–510, Nov. 2018.
10A. Ianus, D. C. Alexander, H. Zhang, and M. Palombo, “Mapping complex cell morphology in the grey matter with double diffusion encoding MR: A simulation study,” NeuroImage, vol. 241, p. 118424, Nov. 2021.
11I. O. Jelescu, A. de Skowronski, F. Geffroy, M. Palombo, and D. S. Novikov, “Neurite Exchange Imaging (NEXI): A minimal model of diffusion in gray matter with inter-compartment water exchange,” NeuroImage, vol. 256, p. 119277, Aug. 2022.
Figures