2372
Longitudinal Dynamic fMRI Analysis of Music vs Singing Perception in Preterm Infants1Division of Development and Growth, Department of Pediatrics, University of Geneva, Geneva, Switzerland, 2Neuro-X Institute, Ecole Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Switzerland, 3Department of Radiology and Medical Informatics, University of Geneva, Geneva, Switzerland
Synopsis
Keywords: Neuro, Brain Connectivity, Neurodevelopment, fMRI, Music
Motivation: Instrumental music and singing differ due to the vocal component of the sung melody and present distinct neural processing pathways, though its developmental origins remain unclear.
Goal(s): This study explores the early development of music vs singing brain processing.
Approach: Dynamic PPI-CAPs approach was applied to longitudinal fMRI data of preterm infants at 33- and 40-weeks gestational age.
Results: Singing elicited a stronger activation of salience and language-related networks, while music elicited limbic networks. Default-mode network and right temporal gyrus were elicited for both stimuli longitudinally. Visual network is also involved for both stimuli but only at term-equivalent age.
Impact: This study explores the longitudinal early development of neural processing networks in the context of instrumental and vocal music stimuli and shows that preterm infants already present a specialized processing for these different stimuli.
Introduction
Despite similarities, music and singing are perceived differently and present distinct neural processing in the brain(1). Though both involve musicality, singing incorporates a vocal component. The origins of this distinction during neurodevelopment are unclear. Literature has shown newborns can already distinguish speech from music(2), thus the preterm population offers an opportunity to explore the ontogenesis of this distinction. Very preterm infants underwent longitudinal fMRI at 33- and 40-weeks gestational age (GA) while listening to a melody either played by a flute or sung by a female voice. This study aims to explore the neural origins of the music-singing distinction.Methods
Population: 54 very preterm infants at the University Hospital of Geneva, Switzerland, underwent longitudinal MRI at 33- and 40-weeks GA. After removal for excessive motion (frame-wise displacement>0.5mm) and incomplete scans, the cohort comprised 44 preterm infants at 33-weeks and 46 at 40-weeks.MR Acquisition: Newborns underwent MRI at both timepoints on a 3.0T Siemens Magnetom scanner, comprising T2-weighted images (voxel-size 0.8x0.8x1.2mm3) and fMRI images acquired via T2*-weighting with the following parameters: 590 images, 36 slices, TR=700ms, voxel-size=2.5mm3, multi-band factor=4.
Stimuli: The task-based fMRI had 4 conditions, each lasting 8 seconds in pseudo-randomized order: a wordless melody either sung by a female voice or played by an Indian flute, silence, and white noise. Each condition was repeated 5 times in a block-design.
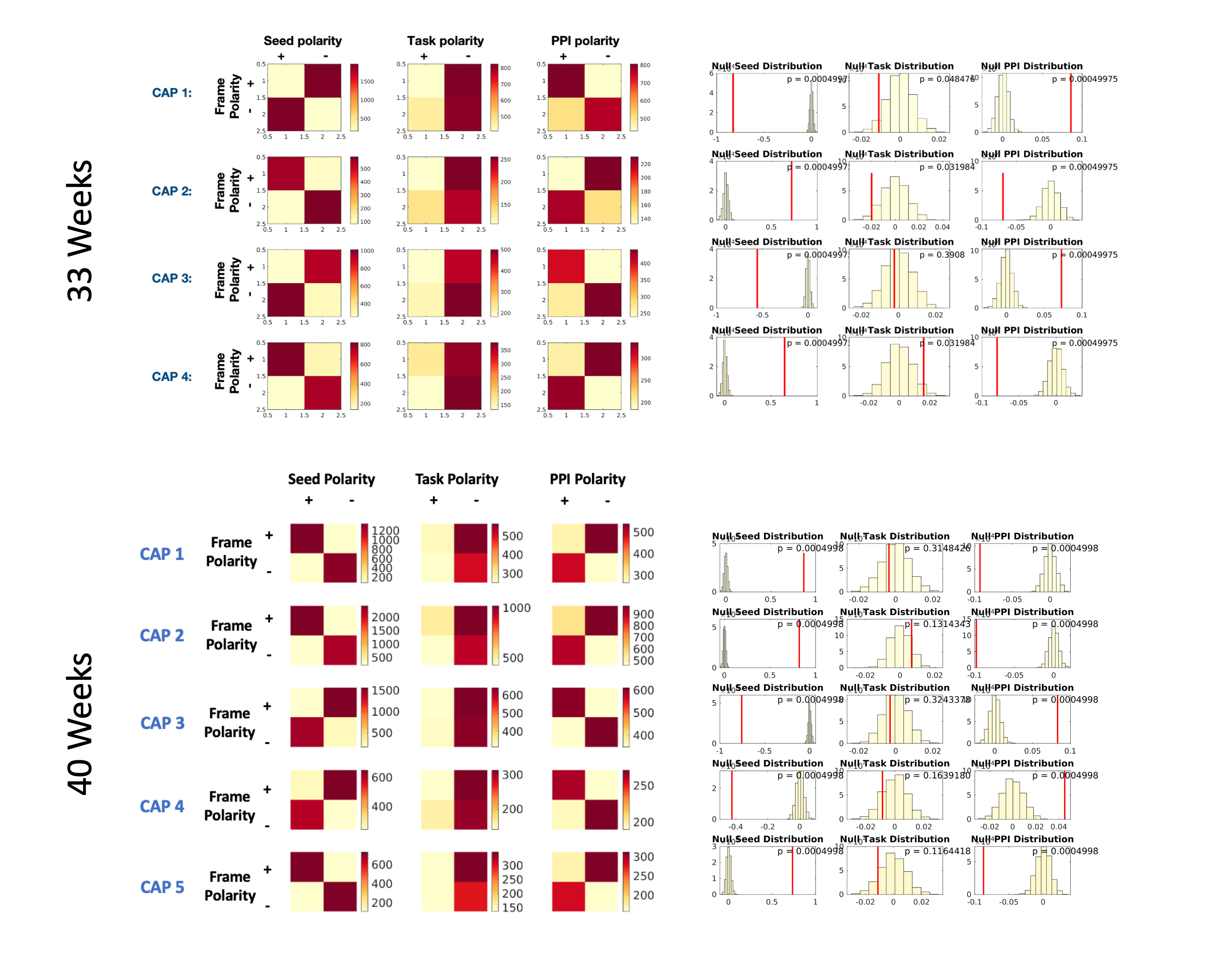
Data Analysis: We employed psychophysiological interaction of co-activation patterns (PPI-CAPs) to explore dynamic, task-based changes in functional connectivity during fMRI acquisition(3). This approach selects moments when a seed region (auditory cortex) is highly active, and clusters frames based on their co-activation patterns (CAPs). Each CAP is then tested whether it varies according to the seed activity, task, or interaction between the two (PPI-effect). The number of CAPs was determined using consensus clustering (k=4 for 33-weeks, k=5 for 40-weeks).
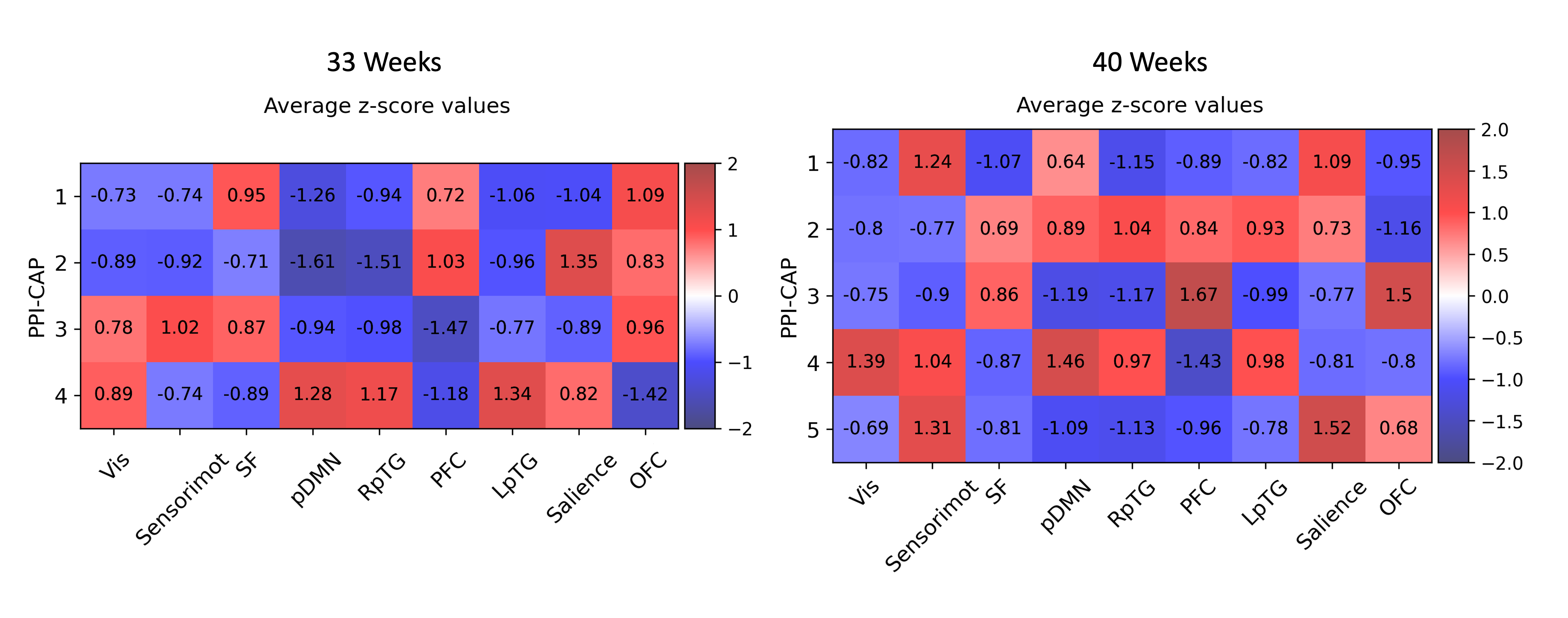
Network Assignment: To assign relevance to identified CAPs, we overlayed ICA-derived functional networks from a previous study(4) with the following regions: visual, sensorimotor, superior frontal (SF), posterior default mode network (combined posterior cingulate cortex and precuneus; pDMN), right posterior temporal gyrus (RpTG), prefrontal (PFC), left posterior temporal gyrus (LpTG), salience, orbitofrontal (OFC).
Results
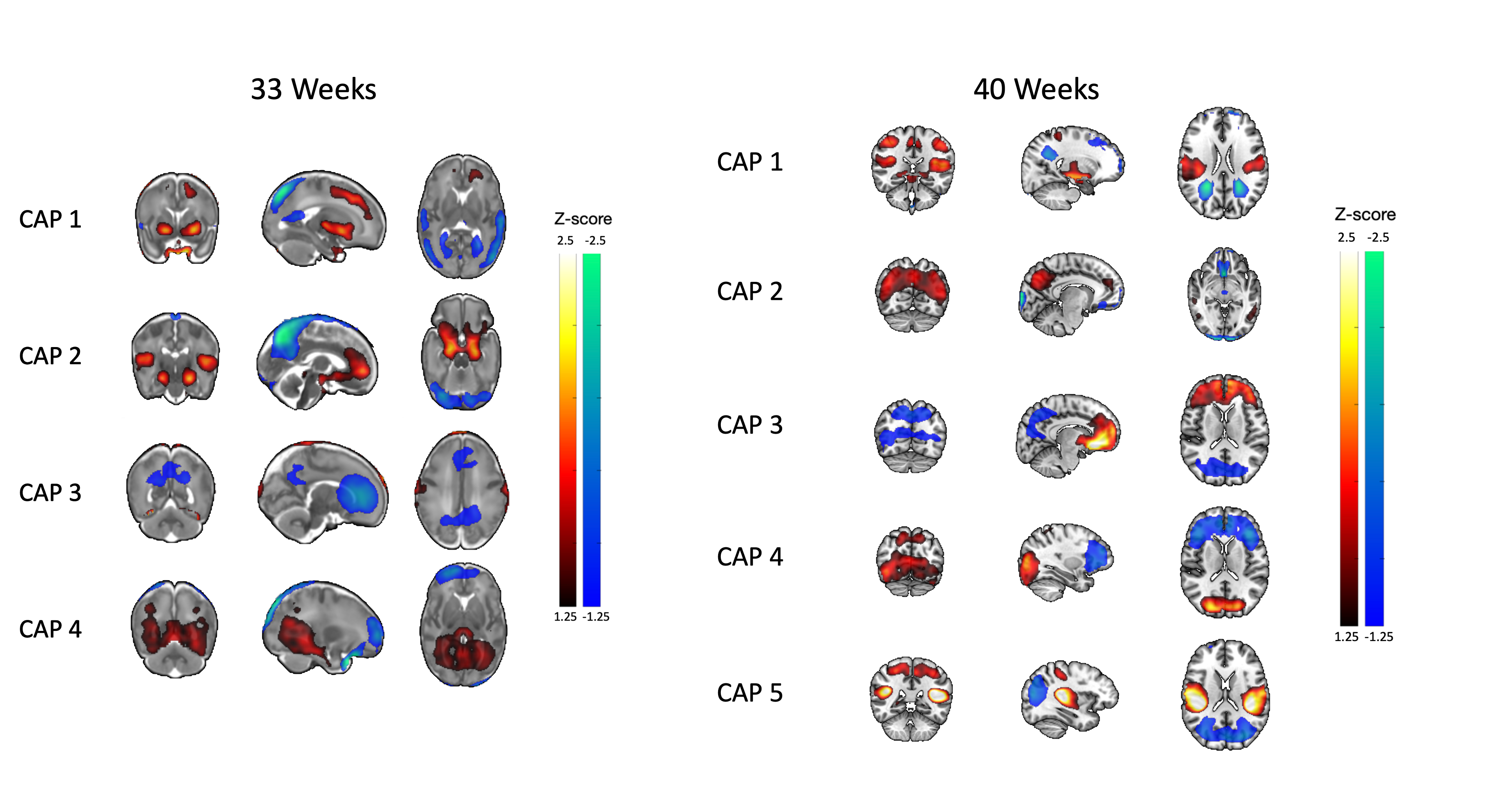
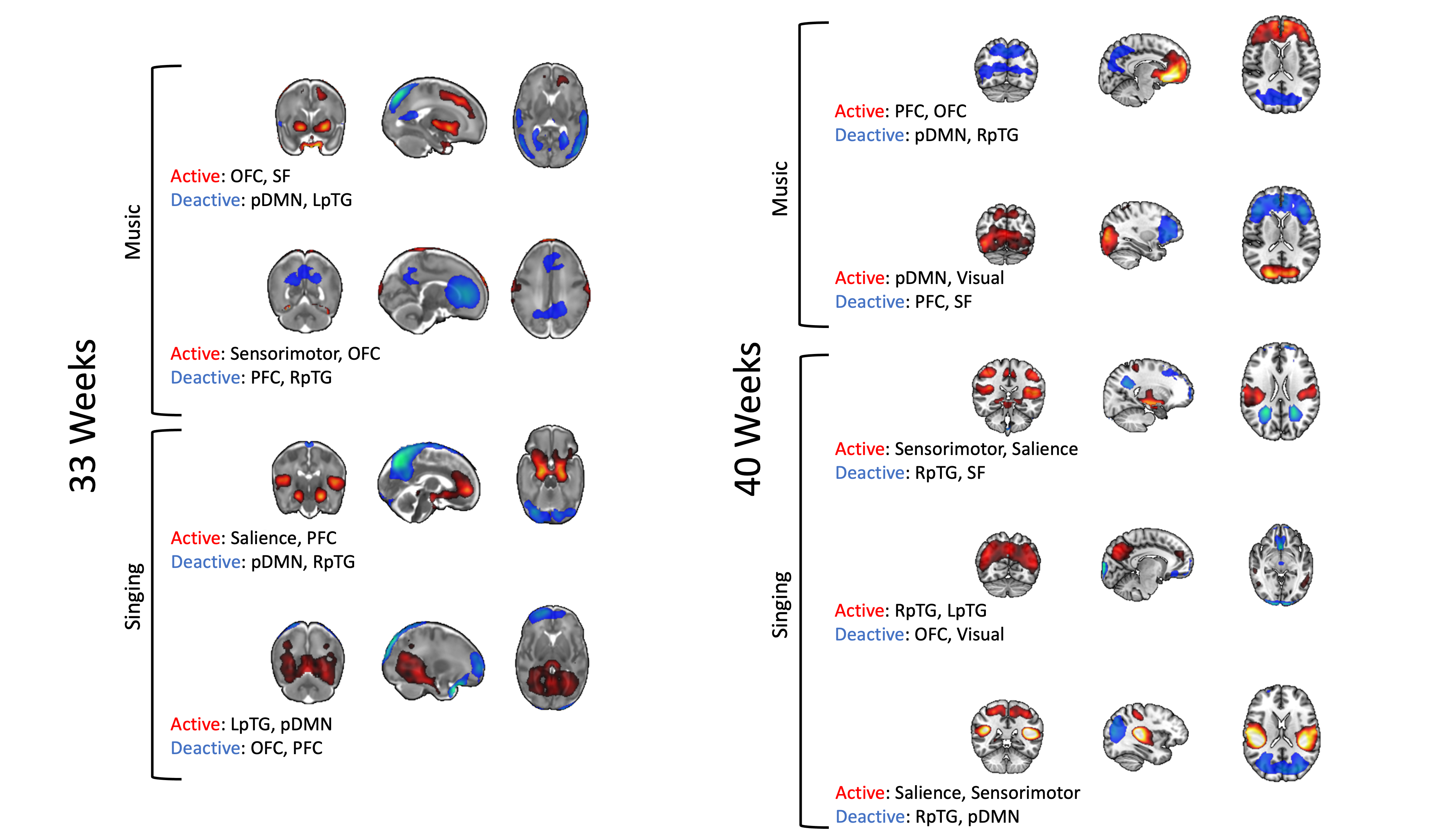
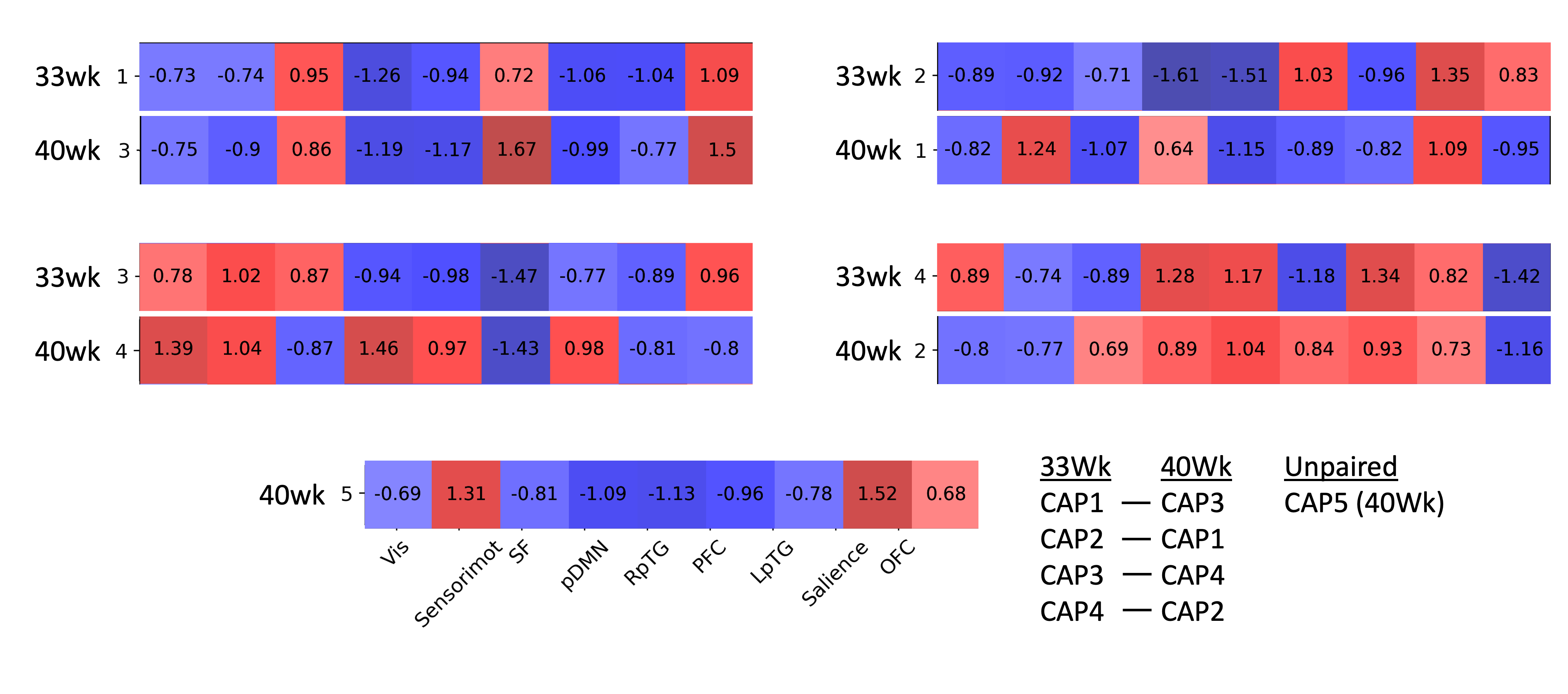
All CAPs showed significant PPI-effect (p<0.0005), demonstrating seed-task interaction (Figure 1). CAPs 1 and 3 of 33-weeks and 3 and 4 of 40-weeks had positive PPI-effect, correlating with the seed during music, while CAPs 2 and 4 of 33-weeks and 1, 2, and 5 of 40-weeks correlated with singing. The dynamic PPI-CAPs approach allowed us to determine the two most active and two most deactive regions that are co-active with the seed based on Z-scores for vocal and instrumental music processing (Figures 2 and 3). A summary of results is shown in Figure 4.The Hungarian algorithm was applied to quantitatively pair the most similar CAPs between 33- and 40-weeks (Figure 5).Discussion/Conclusion
During singing at both timepoints we observe activation of the salience network absent in music, confirming salience of the vocal stimulus in newborns, and of the LpTG that evokes early language pathways, as LpTG activation has shown involvement in speech perception/production later in life(5). At 40, but not 33-weeks, we observe activation of the sensorimotor network for singing but not instrumental music, consistent with literature highlighting the specific role of the audio-articulatory loop during voice perception(6).At both timepoints, during instrumental music we observe OFC activation, absent from singing. The OFC has been linked to emotional and reward processing of stimuli(7) and has consistently shown activation by music listening(8,9,10). In addition, the OFC has been shown to undergo cortical maturation in preterm infants exposed to an early instrumental music intervention(11). This finding provides evidence that music activates limbic networks already early in life. Regions common across co-activation patterns include pDMN and RpTG, involved in cognition and regulation of attentional states(12,13,14), and auditory processing(15), respectively.The visual network activates at 40-weeks for music, while deactivates for singing, and does not contribute at 33-weeks. This suggests that there is no multisensory response at 33-weeks, which is later present during the non-salient instrumental music stimulus at 40-weeks.
Correlation of PPI-CAPs with the Hungarian algorithm, left CAP 5 unpaired, suggesting this co-activation pattern may be “new” and not yet present at 33-weeks. This CAP involves co-activation of salience and sensorimotor regions, suggesting increased salience to auditory stimuli at 40-weeks.
These results provide evidence of neural processing differences in instrumental music and singing perception as early as 33-weeks GA and contribute to better understanding the specialized early development of musical auditory processing for both instrumental and vocal stimuli.
Acknowledgements
No acknowledgement found.References
1. Whitehead JC, Armony JL. Singing in the brain: Neural representation of music and voice as revealed by fMRI. Human brain mapping. 2018;39(12):4913-24.2. Dehaene-Lambertz G, Montavont A, Jobert A, Allirol L, Dubois J, Hertz-Pannier L, et al. Language or music, mother or Mozart? Structural and environmental influences on infants’ language networks. Brain and language. 2010;114(2):53-65.
3. Freitas LG, Bolton TA, Krikler BE, Jochaut D, Giraud A-L, Hüppi PS, et al. Time-resolved effective connectivity in task fMRI: Psychophysiological interactions of Co-Activation patterns. NeuroImage. 2020;212:116635.
4. Lordier L, Meskaldji D-E, Grouiller F, Pittet MP, Vollenweider A, Vasung L, et al. Music in premature infants enhances high-level cognitive brain networks. Proceedings of the National Academy of Sciences. 2019;116(24):12103-8.
5. Buchsbaum, Bradley & Hickok, Gregory & Humphries, Colin. (2001). Role of left posterior superior temporal gyrus in phonological processing for speech perception and production. Cognitive Science. 25. 663-678. 10.1016/S0364-0213(01)00048-9.
6. Lévêque Y, Schön D. Listening to the human voice alters sensorimotor brain rhythms. PloS one. 2013;8(11):e80659.
7. Kam JWY, Solbakk AK, Funderud I, Endestad T, Meling TR, Knight RT. Orbitofrontal damage reduces auditory sensory response in humans. Cortex. 2018 Apr;101:309-312.
8. Koelsch S. Towards a neural basis of music-evoked emotions. Trends Cogn Sci. 2010 Mar;14(3):131-7.
9. Koelsch S. Brain correlates of music-evoked emotions. Nat Rev Neurosci. 2014 Mar;15(3):170-80.
10. Koelsch S. Investigating the Neural Encoding of Emotion with Music. Neuron. 2018 Jun 27;98(6):1075-1079.
11. Sa de Almeida J, Baud O, Fau S, Barcos-Munoz F, Courvoisier S, Lordier L, Lazeyras F, Hüppi PS. Music impacts brain cortical microstructural maturation in very preterm infants: A longitudinal diffusion MR imaging study. Dev Cogn Neurosci. 2023 Jun;61:101254.
12. Mars RB, Neubert FX, Noonan MP, Sallet J, Toni I, Rushworth MF. On the relationship between the "default mode network" and the "social brain". Front Hum Neurosci. 2012 Jun 21;6:189.
13. Leech R, Sharp DJ. The role of the posterior cingulate cortex in cognition and disease. Brain. 2014 Jan;137(Pt 1):12-32.
14. McKiernan KA, Kaufman JN, Kucera-Thompson J, Binder JR. A parametric manipulation of factors affecting task-induced deactivation in functional neuroimaging. J Cogn Neurosci. 2003 Apr 1;15(3):394-408.
15. Ohnishi T, Matsuda H, Asada T, Aruga M, Hirakata M, Nishikawa M, Katoh A, Imabayashi E. Functional anatomy of musical perception in musicians. Cereb Cortex. 2001 Aug;11(8):754-60.
Figures