2371
Microstructural cortical maturation underlies longitudinal BOLD signal variability of emerging resting-state networks in preterm infants1Department of Pediatrics, HUG, Geneva University Hospital, University of Geneva, Geneva, Switzerland, 2University of Geneva, Geneva, Switzerland, 3Center of Biomedical Imaging (CIBM), University of Geneva, Geneva, Switzerland, 4Neuro-X-Institute, Ecole Polytechnique Federal de Lausanne (EPFL), Lausanne, Switzerland
Synopsis
Keywords: Neonatal, Normal development, early preterm brain development
Motivation: BOLD signal variability (BOLD-SD) has emerged as a measure for assessing brain function, but little is known regarding its biological significance.
Goal(s): Demonstrate that cortical BOLD-SD modifications are accompanied by structural intracortical maturational changes. Elucidate brain networks undergoing the most important maturational changes during early development.
Approach: Longitudinal brain MRI acquisition in preterm infants at 33 and 40 weeks’ gestational age. Assessment of cortical BOLD-SD and NODDI indices longitudinal modifications per brain network.
Results: A significant longitudinal cortical BOLD-SD increase is observed in primary sensory networks and Default-Mode-Network, accompanied by a decreased NDI (neurite-density-index) and/or increased ODI (orientation-dispersion-index), reflecting concomitant structural intracortical maturation.
Impact: During early brain development, the BOLD signal variability increase in resting-state networks was associated to underlying structural intracortical maturational changes and thus it can be considered as a marker of cortical maturation.
Introduction
Resting-state functional MRI (RS-fMRI) measures fluctuations of the blood oxygenation level-dependent (BOLD) signal intensity over time as a consequence of oxygenated-blood-flow supplying active neurons (1). BOLD signal variability (BOLD-SD, calculated as the standard-deviation (SD) of BOLD signal) has been shown to predict cognitive performance and functional integrity (2, 3) and has been used as a relevant measure when examining age-related changes in brain function (4, 5).During preterm infants’ development, regional and age-specific RS-fMRI cortical activation patterns are observed (6, 7), but little is known regarding BOLD-SD and concomitant cortical structural maturation. We hypothesize that early BOLD-SD modifications might reflect underlying structural developmental changes and thus cortical maturation.We aim to compare functional spatio-temporal changes of RS-fMRI BOLD-SD with concomitant cortical structural changes of relevant RS-networks during preterm infants’ early brain development.Methods
Population: 54 very-preterm infants were recruited at Geneva University Hospital, Switzerland, and underwent a longitudinal MRI-scan at time-points 33 and 40 weeks’ gestational age (GA).MRI acquisition at both time-points was performed on a 3.0T MR-scanner Siemens Magnetom comprising: T2-weighted images (voxel size 0.8 × 0.8 × 1.2 mm3), multi-shell diffusion imaging (MSDI, voxel size 1.8 mm3, 4 b0, 10 non-collinear directions b= 200 s/mm2, 30 b= 1000 s/mm2, 50 b= 2000 s/mm2) and RS-fMRI of 7 minutes’ duration (voxel size 2.5mm3).
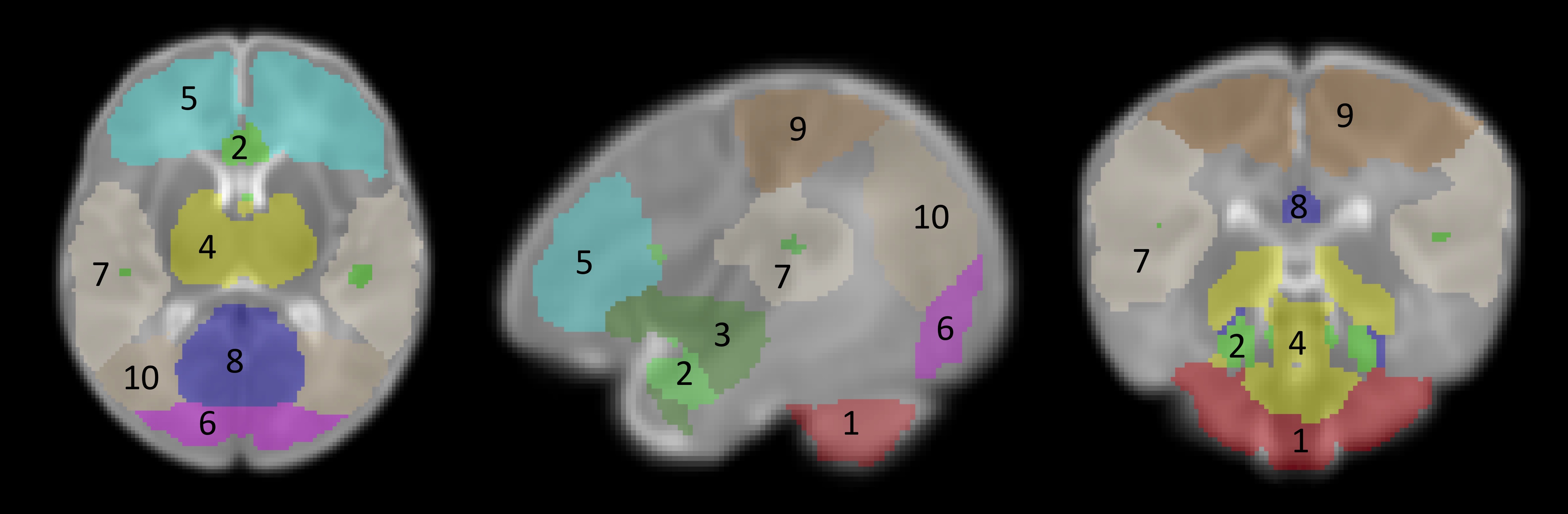
Atlas construction: a RS-fMRI brain atlas based on ICA (independent component analysis) components comprising 10 brain networks was created from our data set (Figure 1).
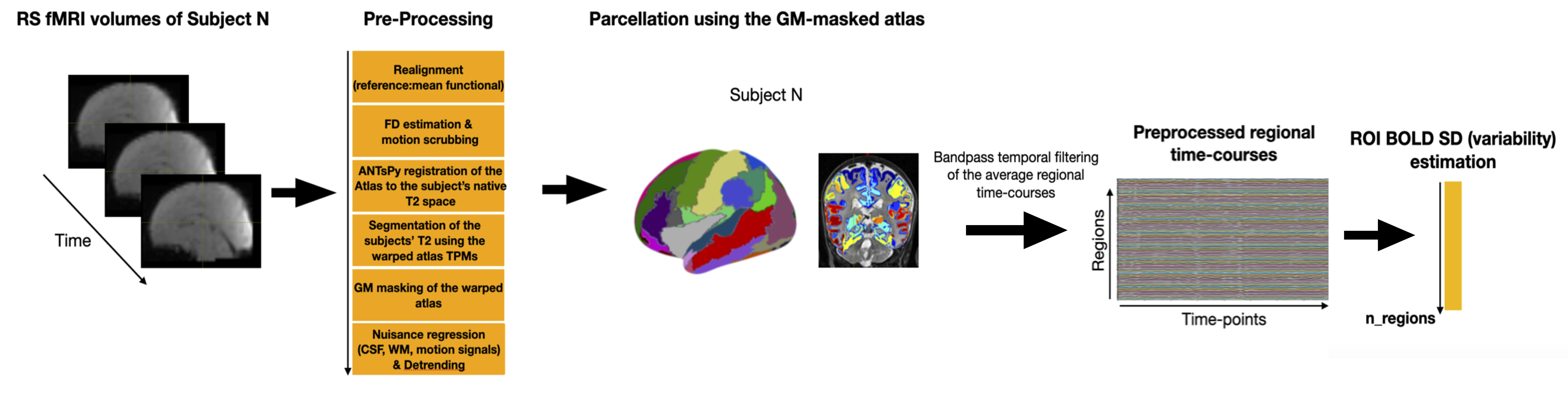
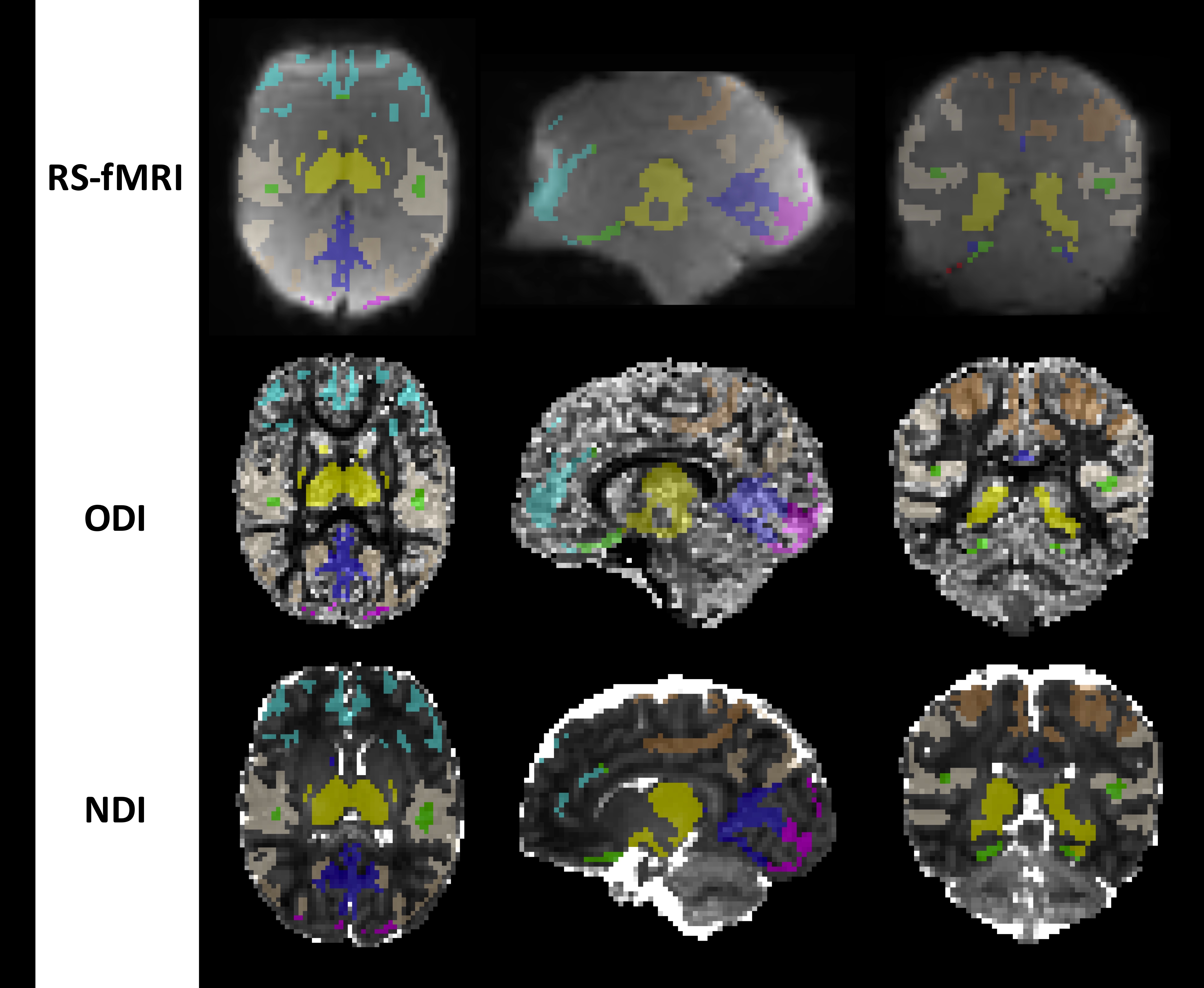
RS-fMRI data processing was conducted using the pipeline described in Figure 2. The ICA atlas was registered to each subject’s space using ANTs (8). Average regional BOLD time-courses were extracted for each subject and BOLD-SD was estimated at each time-point per brain region in grey-matter (GM) voxels (Figure 2 and 3). Final sample comprised 29 subjects.
MSDI data was preprocessed using MRtrix3 (9) and analyzed by means of NODDI model (10) using Bingham-distribution for ODI (orientation dispersion index) and NDI (neurite density index) estimation. The ICA atlas was registered to each subject’ space using ANTs (8). Only GM voxels mean values were extracted and averaged per brain region (Figure 3). The final sample comprised 39 subjects.
Statistical analysis: mean BOLD-SD, ODI and NDI, per brain region, were evaluated for each subject longitudinally. Paired Samples t-tests were conducted to determine significant differences. False discovery rate (FDR) was used for multiple comparisons correction at α=0.05. Multiple regression analysis was used to evaluate the relationship between BOLD-SD delta (variation from 33 to 40 weeks GA) and the predictors: ODI delta and NDI delta.
Results
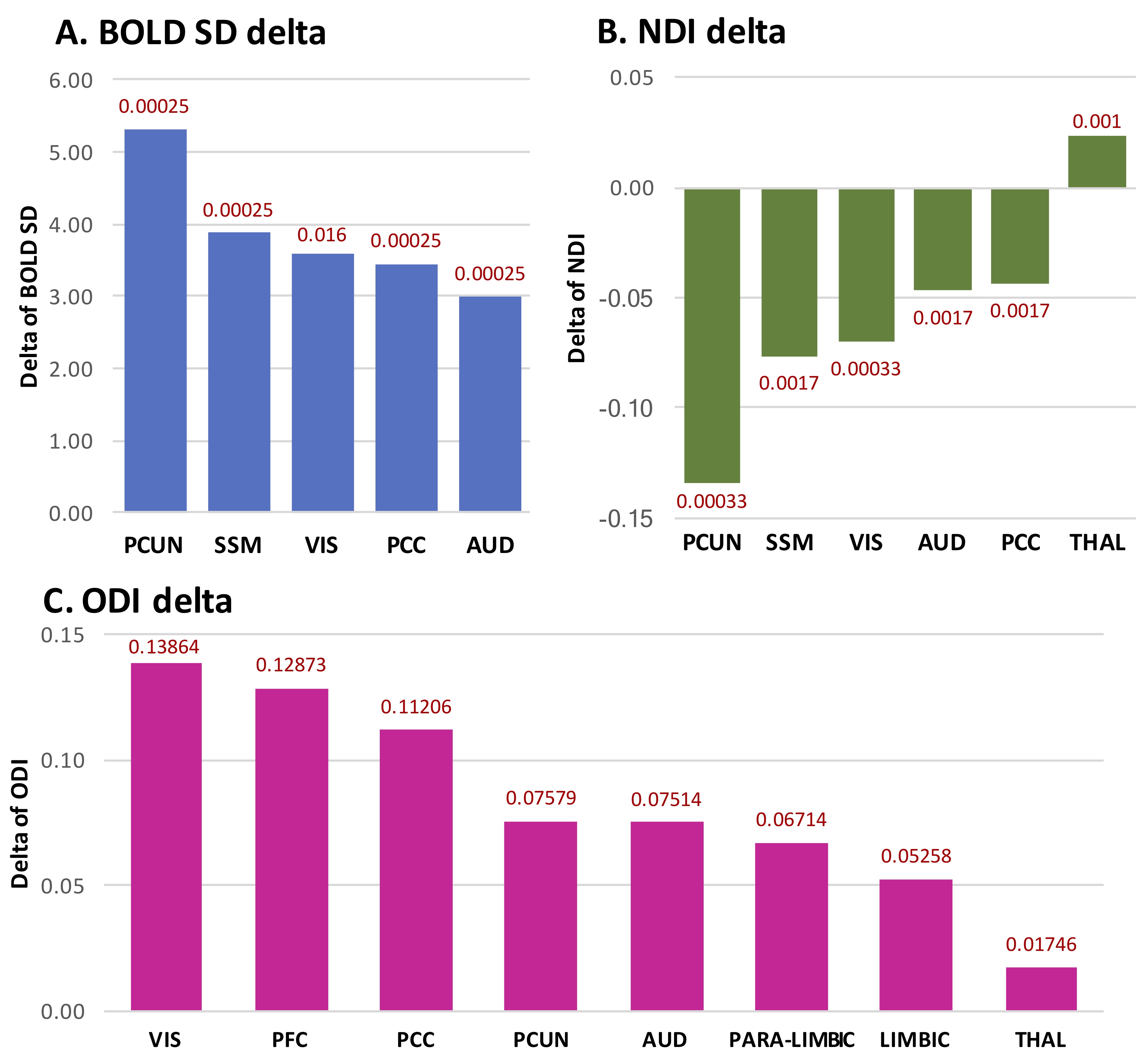
From 33 to 40 weeks GA, BOLD-SD significantly increased in 5 brain regions, in descending significance order: precuneus, sensorimotor, visual, posterior cingulate gyrus (PCC) and auditory regions (Figure 4A). Regarding NODDI cortical changes, NDI decreased significantly in 6 regions, in descending significance order: precuneus, sensorimotor, visual, auditory and PCC; whereas it increased significantly in the thalamus/brainstem (Figure 4B). ODI increased significantly in 8 regions, in descending significance order: visual, PFC, PCC, precuneus, auditory, paralimbic-belt, limbic and thalamus/brainstem regions (Figure 4C). NDI delta significantly predicted BOLD-SD delta in PFC (p = 0.043) and PCC (p = 0.035), being marginally significant as a predictor of BOLD-SD in sensorimotor cortex (p = 0.073). ODI delta significantly predicted BOLD-SD delta in sensorimotor cortex (p = 0.013).Discussion/Conclusion
From 33 to 40 weeks GA, BOLD-SD increased significantly in sensorimotor, visual, auditory, precuneus and PCC regions. Sensorimotor, visual and auditory are responsible for primary-sensory processing. A longitudinal BOLD-SD increase in these regions is in line with the establishment of sensory-thalamo-cortical-connectivity and thus related cortical functional modifications. The precuneus and PCC constitute together the posterior part of the “default-mode-network” (pDMN), a fundamental brain system functionally linked to self-related processes (11, 12) and attention/cognitive processing (13, 14). This finding supports the emergence of DMN in this early period of development, in agreement with recent literature (15).The BOLD-SD increase was accompanied by an underlying longitudinal structural cortical maturation proven by an increased ODI and/or decreased NDI. The observed intracortical ODI increase and NDI decrease until term-age is in agreement with previous literature and has been related to increased dendritic arborisation and cortical complexity (16, 17). Of notice, the magnitude of the BOLD-SD increase in the visual, auditory, precuneus and PCC regions was parallel to the NDI decrease. Furthermore, NDI decrease was shown to predict BOLD-SD increase in PCC and PFC, being marginally significant for sensorimotor cortex, while ODI increase was shown to predict BOLD-SD increase in sensorimotor cortex. Our findings suggest thus that BOLD-SD might be used as a marker of cortical maturation.Acknowledgements
The authors thank all clinical staff, namely in Neonatology and Unit of Development of the Children’s Hospital, HUG, all parents and newborns participating in the project, the Pediatrics Clinic Research Platform and the Center for Biomedical Imaging (CIBM) of the University Hospitals of Geneva, for all their valuable help and support. This study was supported by grants from the Swiss National Science Foundation (no. 32473B_135817/1 and no. 324730–163084), the Prim'enfance Foundation, the Swiss Government Excellence Scholarship (no. 2017.0450/OP), the Swiss Academy of Medical Sciences (YTCR 49/19) and the Fondation pour la recherche en périnatalité.
References
1. Buxton RB, Uludag K, Dubowitz DJ, Liu TT. Modeling the hemodynamic response to brain activation. Neuroimage. 2004;23:S220-S33. doi: 10.1016/j.neuroimage.2004.07.013. PubMed PMID: WOS:000225374100021.
2. Protzner AB, Kovacevic N, Cohn M, McAndrews MP. Characterizing Functional Integrity: Intraindividual Brain Signal Variability Predicts Memory Performance in Patients with Medial Temporal Lobe Epilepsy. Journal of Neuroscience. 2013;33(23):9855-65. doi: 10.1523/Jneurosci.3009-12.2013. PubMed PMID: WOS:000319963300029.
3. Roberts R.P. WK, Moreau D., Addis D.R. Reassessing the Functional Significance of BOLD Variability. bioRxiv. 2023. doi: doi: https://doi.org/10.1101/2023.02.06.527384.
4. Garrett DD, Kovacevic N, McIntosh AR, Grady CL. The Modulation of BOLD Variability between Cognitive States Varies by Age and Processing Speed. Cerebral Cortex. 2013;23(3):684-93. doi: 10.1093/cercor/bhs055. PubMed PMID: WOS:000316275400018.
5. Grady CL, Garrett DD. Understanding variability in the BOLD signal and why it matters for aging. Brain Imaging and Behavior. 2014;8(2):274-83. doi: 10.1007/s11682-013-9253-0. PubMed PMID: WOS:000335765700010.
6. Smyser CD, Inder TE, Shimony JS, Hill JE, Degnan AJ, Snyder AZ, Neil JJ. Longitudinal analysis of neural network development in preterm infants. Cereb Cortex. 2010;20(12):2852-62. doi: 10.1093/cercor/bhq035. PubMed PMID: 20237243; PMCID: PMC2978240.
7. Doria V, Beckmann CF, Arichi T, Merchant N, Groppo M, Turkheimer FE, Counsell SJ, Murgasova M, Aljabar P, Nunes RG, Larkman DJ, Rees G, Edwards AD. Emergence of resting state networks in the preterm human brain. Proc Natl Acad Sci U S A. 2010;107(46):20015-20. doi: 10.1073/pnas.1007921107. PubMed PMID: 21041625; PMCID: PMC2993415.
8. Avants BB, Tustison NJ, Song G, Cook PA, Klein A, Gee JC. A reproducible evaluation of ANTs similarity metric performance in brain image registration. Neuroimage. 2011;54(3):2033-44. doi: 10.1016/j.neuroimage.2010.09.025. PubMed PMID: WOS:000286302000027.
9. Tournier JD, Smith R, Raffelt D, Tabbara R, Dhollander T, Pietsch M, Christiaens D, Jeurissen B, Yeh CH, Connelly A. MRtrix3: A fast, flexible and open software framework for medical image processing and visualisation. Neuroimage. 2019;202:116137. doi: 10.1016/j.neuroimage.2019.116137. PubMed PMID: 31473352.
10. Zhang H, Schneider T, Wheeler-Kingshott CA, Alexander DC. NODDI: Practical in vivo neurite orientation dispersion and density imaging of the human brain. Neuroimage. 2012;61(4):1000-16. doi: 10.1016/j.neuroimage.2012.03.072. PubMed PMID: WOS:000305920600028.
11. Cavanna AE, Trimble MR. The precuneus: a review of its functional anatomy and behavioural correlates. Brain. 2006;129:564-83. doi: 10.1093/brain/awl004. PubMed PMID: WOS:000235433800003.
12. Fransson P, Marrelec G. The precuneus/posterior cingulate cortex plays a pivotal role in the default mode network: Evidence from a partial correlation network analysis. Neuroimage. 2008;42(3):1178-84. doi: 10.1016/j.neuroimage.2008.05.059. PubMed PMID: WOS:000258864800012.
13. Fair DA, Cohen AL, Dosenbach NU, Church JA, Miezin FM, Barch DM, Raichle ME, Petersen SE, Schlaggar BL. The maturing architecture of the brain's default network. Proc Natl Acad Sci U S A. 2008;105(10):4028-32. doi: 10.1073/pnas.0800376105. PubMed PMID: 18322013; PMCID: PMC2268790.
14. Whitfield-Gabrieli S, Ford JM. Default Mode Network Activity and Connectivity in Psychopathology. Annual Review of Clinical Psychology, Vol 8. 2012;8:49-+. doi: 10.1146/annurev-clinpsy-032511-143049. PubMed PMID: WOS:000304200200004.
15. Yu QL, Ouyang MH, Detre J, Kang HY, Hu D, Hong B, Fang F, Peng Y, Huang H. Infant brain regional cerebral blood flow increases supporting emergence of the default-mode network. Elife. 2023;12. doi: ARTN e78397 10.7554/eLife.78397. PubMed PMID: WOS:000920203400001.
16. Batalle D, O'Muircheartaigh J, Makropoulos A, Kelly CJ, Dimitrova R, Hughes EJ, Hajnal JV, Zhang H, Alexander DC, Edwards AD, Counsell SJ. Different patterns of cortical maturation before and after 38 weeks gestational age demonstrated by diffusion MRI in vivo. Neuroimage. 2019;185:764-75. doi: 10.1016/j.neuroimage.2018.05.046. PubMed PMID: WOS:000451628200064.
17. Sa de Almeida J, Baud O, Fau S, Barcos-Munoz F, Courvoisier S, Lordier L, Lazeyras F, Huppi PS. Music impacts brain cortical microstructural maturation in very preterm infants: A longitudinal diffusion MR imaging study. Dev Cogn Neuros-Neth. 2023;61. doi: ARTN 101254 10.1016/j.dcn.2023.101254. PubMed PMID: WOS:001002048300001.
Figures