1861
Unlocking Myelin Mysteries: Could 31-Phosphorous Solid-State NMR be the key to quantitative MRI of myelin membrane morphology?1Physics and Astronomy, The University of British Columbia, Vancouver, BC, Canada, 2International Collaboration on Repair Discoveries (ICORD), The University of British Columbia, Vancouver, BC, Canada, 3Radiology, The University of British Columbia, Vancouver, BC, Canada, 4Pathology and Laboratory Medicine, The University of British Columbia, Vancouver, BC, Canada, 5UBC MRI Research Centre, The University of British Columbia, Vancouver, BC, Canada
Synopsis
Keywords: Non-Proton, White Matter, Myelin
Motivation: Improved sensitivity to changes in myelin membranes may be achieved by phosphorous (31P) solid-state NMR (ssNMR), enabling more direct evaluation of neurodegeneration.
Goal(s): To demonstrate how ssNMR proton cross-polarization (CP) may be more sensitive to membrane morphology than 31P alone.
Approach: Using porcine neural tissue, we conducted a series of 31P ssNMR experiments that characterize the myelin phospholipid involvement in 31P-CP and CP’s sensitivity to variations in membrane composition, orientation, and dynamics.
Results: The CP signal is highly sensitive to the amount and orientation of myelin between grey and white matter samples across neural regions, and may better detect changes in membrane structure.
Impact: Improved sensitivity to subtle variations in myelin membrane morphology using the 31P ssNMR method of CP has the potential for in vivo MRI use, and could lead to earlier diagnosis, as well as enhanced disease and treatment monitoring.
INTRODUCTION
Novel approaches to investigate myelin integrity are urgently needed. Despite being an active area of research1-3, existing myelin imaging metrics rely on relatively indirect measures of myelin lipids.Previous work using the solid-state NMR (ssNMR) technique of cross-polarization (CP)4,5 between phosphorous (31P) and protons (1H) in phospholipids selectively probed myelin membranes. The potential for translation of 1H/31P CP to in vivo MRI was also demonstrated by pairing magnetization transfer (MT) with CP, generating a hybrid MT-CP sequence capable of shuttling polarization between aqueous 1H and myelin 31P6,7.
31P ssNMR line shapes in lipid bilayers reflect membrane composition and structure, and are sensitive to variations in phospholipid orientation, organization, and dynamics8-16. Our objective was to investigate myelin phospholipid membranes using 31P ssNMR to characterize microstructural information about these membranes.
METHODS
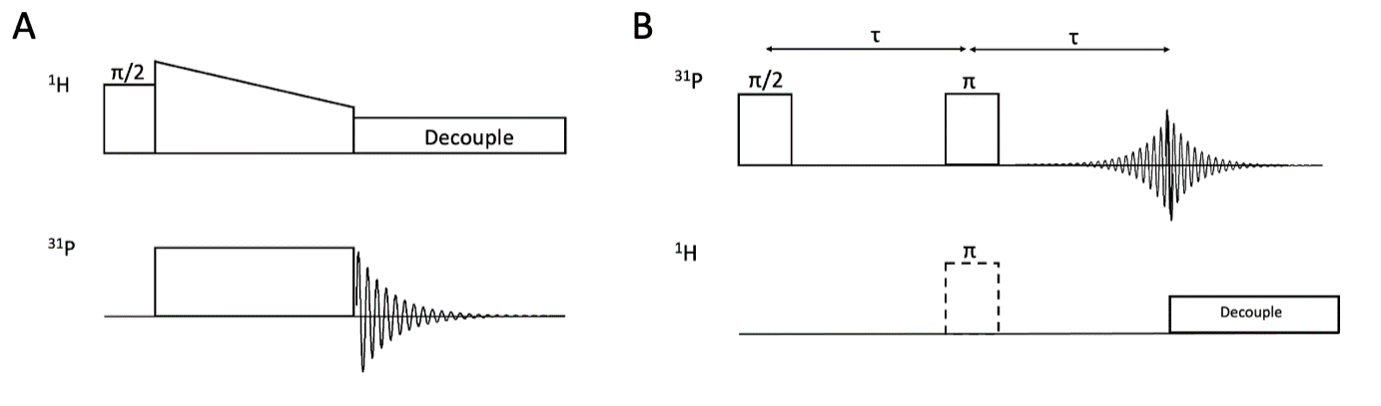
Experiments included a Ramped Amplitude CP17 and a Spin-Echo DOuble Resonance (SEDOR)18 (Fig.1) on a 8.4T ssNMR spectrometer19 (horizontal coil double-resonance probe, 21oC). Fresh porcine samples included cortical gray matter (GM) and white matter (WM, brain stem (WM1), corpus callosum (WM2), pons (WM3), spinal cord).CP membrane sensitivity experiments were performed at 9.4T (Varian INOVA spectrometer, Varian/Chemagnetics HXY-T1 MAS probe) at -20°C, -25°C, -30°C. Spinal cord WM was compared to corresponding 21°C controls from the same spinal cord taken on the 8.4T spectrometer.
RESULTS
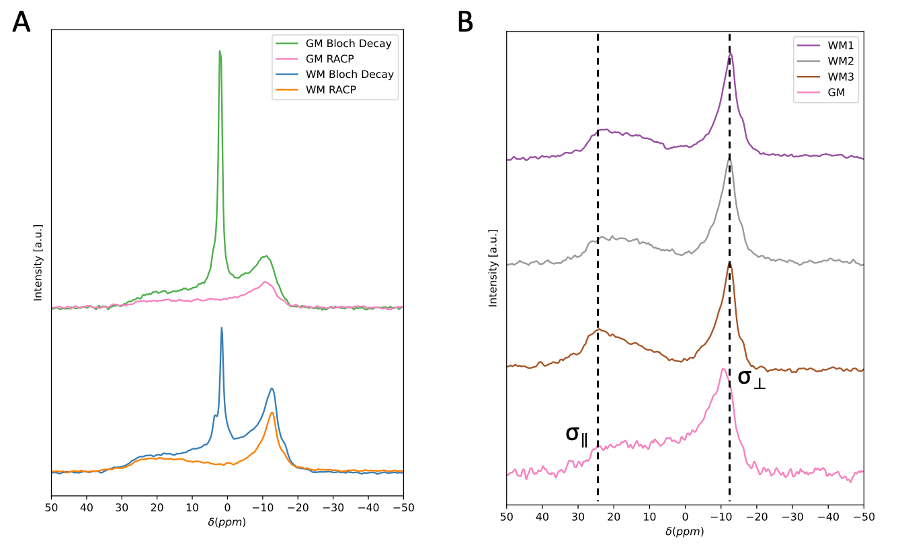
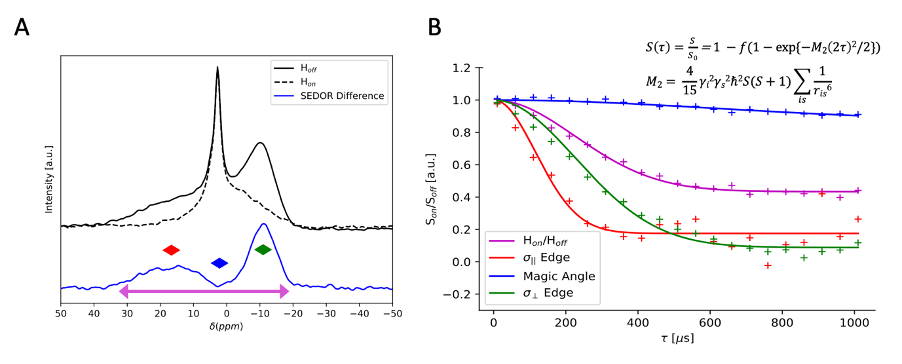
Fig.2 compares 31P-CP line shapes and efficiency across neural regions. The standard 31P ssNMR spectra of GM and WM exhibit a sharp aqueous peak from inorganic phosphates superimposed on characteristically broad powder patterns from phospholipid membranes. Both CP spectra exhibit aqueous peak filtering. GM and WM 31P spectra show pronounced differences in phospholipid membrane contribution to the overall spectrum, which are retained in their 31P-CP spectra. Comparing GM CP spectra to different WM regions shows signal sensitivity to the degree of membrane orientation. WM spectra exhibit a dip at the magic angle where dipolar couplings vanish and a higher enhancement at the parallel edge where couplings are strongest, neither of which are resolvable in GM.Fig.3 summarizes spinal cord WM SEDOR experiments. Dipolar modulated 31P spectrum indicates coaxial alignment of the dipolar coupling and 31P chemical shift tensors, along the phospholipid molecule. Second moment (M2) analysis at the σ|| edge reveals an effective 1H-31P bond length of ~2.93Å, generating a coupling strength of ~1900Hz, roughly double that of the perpendicular edge and consistent with the observed orientation dependence in CP spectra.
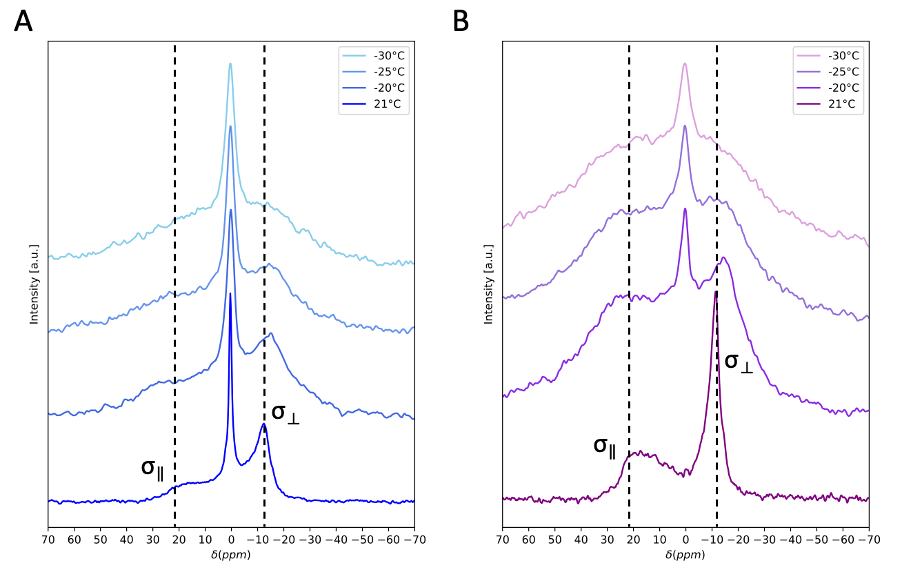
Fig.4 shows spinal cord WM temperature-varied CP experiments. With decreasing temperature, the 31P powder pattern is broadened, and a peak appears in the CP line shape at the position of the aqueous peak, both consistent with a reduction in molecular motions which reduce averaging of 1H-31P dipolar couplings. Between -25oC and -30oC differences in the line shape between the CP and 31P spectra appear to reflect changes in membrane organization and dynamics.
DISCUSSION
Variations in line shapes of neural 31P-CP spectra indicate sensitivity to differences in myelin abundance and membrane orientation, which is consistent with results from SEDOR-difference experiments indicating the coaxial alignment of the dipolar and 31P chemical shift tensors. Thus, the CP spectrum is sensitive to orientation of the phospholipid headgroup within the membrane, making the net spectrum sensitive to the homogeneity of all myelin contributions.Temperature-varied experiments on WM hint that CP spectra exhibit greater sensitivity to changes in myelin membrane dynamics than 31P-only spectra. This is apparent with the emergence of a sharp peak in the cooled CP spectra, which may arise when tumbling of aqueous 31P metabolites is sufficiently slow so as to not average the dipolar couplings necessary for CP. The CP spectra also demonstrate sensitivity to changes in myelin organization and molecular dynamics. This could indicate that CP is more sensitive to subtle changes in membrane organization. At sufficiently cold temperatures, it is possible that phase polymorphisms in lipid membranes are seeded as water trapped between myelin bilayers crystalizes and disrupts local phospholipid ordering20,21. Such disruptions could result in a mixture of lamellar and inverted hexagonal phospholipid phases, potentially discernable by CP due to its high sensitivity to orientation of phospholipid headgroups within membranes13,22,23.
CONCLUSION
Our results hint towards CP being characteristically more sensitive to subtle changes in phospholipid membrane conformation and dynamics than 31P-only metrics. If true, CP would uniquely poise to detect changes to myelin membranes which may precede demyelinating disease processes and may enable superior diagnosis and monitoring of neurodegenerative diseases.Acknowledgements
This work was conducted on the traditional, ancestral, and unceded territories of Coast Salish Peoples, including the territories of the xwməθkwəy̓əm (Musqueam), Skwxwú7mesh (Squamish), Stó:lō and Səl̓ílwətaʔ/Selilwitulh (Tsleil- Waututh) Nations.
CK, CM, AM and CL gratefully acknowledge funding from the NSERC through CGS-M and Discovery grants programs. AE extends appreciation to MS Canada for a Doctoral Studentship Award.
References
1. Piredda, G. F.; Hilbert, T.; Thiran, J.; Kober, T. Probing Myelin Content of the Human Brain with MRI: A Review. Magn. Reson. Med. 2021, 85 (2), 627–652. https://doi.org/10.1002/mrm.28509.
2. van der Weijden, C. W. J.; Biondetti, E.; Gutmann, I. W.; Dijkstra, H.; McKerchar, R.; de Paula Faria, D.; de Vries, E. F. J.; Meilof, J. F.; Dierckx, R. A. J. O.; Prevost, V. H.; Rauscher, A. Quantitative Myelin Imaging with MRI and PET: An Overview of Techniques and Their Validation Status. Brain 2023, 146 (4), 1243–1266. https://doi.org/10.1093/brain/awac436.
3. Laule, C.; Vavasour, I. M.; Kolind, S. H.; Li, D. K. B.; Traboulsee, T. L.; Moore, G. R. W.; MacKay, A. L. Magnetic Resonance Imaging of Myelin. Neurotherapeutics 2007, 4 (3), 460–484. https://doi.org/10.1016/j.nurt.2007.05.004.
4. Hartmann, S. R.; Hahn, E. L. Nuclear Double Resonance in the Rotating Frame. Phys. Rev. 1962, 128 (5), 2042–2053. https://doi.org/10.1103/PhysRev.128.2042.
5. Pines, A.; Gibby, M. G.; Waugh, J. S. Proton‐Enhanced Nuclear Induction Spectroscopy. A Method for High Resolution NMR of Dilute Spins in Solids. The Journal of Chemical Physics 1972, 56 (4), 1776–1777. https://doi.org/10.1063/1.1677439.
6. Ensworth, A.; Knight, C.-A.; Kozlowski, P.; Laule, C.; MacKay, A. L.; Michal, C. A. 1H-31P Cross-Polarization: A New Frontier to Study Myelin in White Matter.; London, England, 2022.
7. Ensworth, A.; Knight, C.-A.; Kozlowski, P.; Laule, C.; Michal, C. A. Solid State NMR in White Matter: Unconventional 31P→1H Cross Polarization Interrogates the Proton Pool.; Toronto, Canada, 2023.
8. Schiller, J.; Muller, M.; Fuchs, B.; Arnold, K.; Huster, D. 31P NMR Spectroscopy of Phospholipids: From Micelles to Membranes. CAC 2007, 3 (4), 283–301. https://doi.org/10.2174/157341107782109635.
9. Seelig, J.; Borle, F.; Cross, T. A. Magnetic Ordering of Phospholipid Membranes. Biochimica et Biophysica Acta (BBA) - Biomembranes 1985, 814 (1), 195–198. https://doi.org/10.1016/0005-2736(85)90437-7.
10. Watts, A.; Opella, S. J. Membranes Studied by NMR Spectroscopy. In Encyclopedia of Spectroscopy and Spectrometry; Elsevier, 2017; pp 781–789. https://doi.org/10.1016/B978-0-12-803224-4.00201-6.
11. Dufourc, E. J. Solid‐State NMR in Biomembranes. In Chemical Biology; Larijani, B., Rosser, Colin. A., Woscholski, R., Eds.; Wiley, 2006; pp 113–131. https://doi.org/10.1002/9780470319253.ch8.
12. Dufourc, E. J.; Mayer, C.; Stohrer, J.; Althoff, G.; Kothe, G. Dynamics of Phosphate Head Groups in Biomembranes. Comprehensive Analysis Using Phosphorus-31 Nuclear Magnetic Resonance Lineshape and Relaxation Time Measurements. Biophysical Journal 1992, 61 (1), 42–57. https://doi.org/10.1016/S0006-3495(92)81814-3.
13. Klose, G.; Möps, A.; Haage, K. Incorporation of Phosphonic Acid Diesters into Lipid Model Membranes Part I. Influence of the Esters on the Phase Behaviour of Egg Yolk Lecithin Water Systems and Their Orientational Behaviour at Low Water Content as Seen by 2H- and 31P-NMR. Chemistry and Physics of Lipids 1990, 55 (1), 1–11. https://doi.org/10.1016/0009-3084(90)90143-F.
14. Mužić, T.; Tounsi, F.; Madsen, S. B.; Pollakowski, D.; Konrad, M.; Heimburg, T. Melting Transitions in Biomembranes. Biochimica et Biophysica Acta (BBA) - Biomembranes 2019, 1861 (11), 183026. https://doi.org/10.1016/j.bbamem.2019.07.014.
15. Griffin, R. G.; Powers, L.; Pershan, P. S. Head-Group Conformation in Phospholipids: A Phosphorus-31 Nuclear Magnetic Resonance Study of Oriented Monodomain Dipalmitoylphosphatidylcholine Bilayers1".
16. Koivuniemi, A. The Biophysical Properties of Plasmalogens Originating from Their Unique Molecular Architecture. FEBS Letters 2017, 591 (18), 2700–2713. https://doi.org/10.1002/1873-3468.12754.
17. Metz, G.; Wu, X. L.; Smith, S. O. Ramped-Amplitude Cross Polarization in Magic-Angle-Spinning NMR. Journal of Magnetic Resonance, Series A 1994, 110 (2), 219–227. https://doi.org/10.1006/jmra.1994.1208.
18. Kaplan, D. E.; Hahn, E. L. Expériences de double irradiation en résonance magnétique par la méthode d’impulsions. J. Phys. Radium 1958, 19 (11), 821–825. https://doi.org/10.1051/jphysrad:019580019011082100.
19. Michal, C. A.; Broughton, K.; Hansen, E. A High Performance Digital Receiver for Home-Built Nuclear Magnetic Resonance Spectrometers. Review of Scientific Instruments 2002, 73 (2), 453–458. https://doi.org/10.1063/1.1433950.
20. Lee, D.-K.; Kwon, B. S.; Ramamoorthy, A. Freezing Point Depression of Water in Phospholipid Membranes: A Solid-State NMR Study. Langmuir 2008, 24 (23), 13598–13604. https://doi.org/10.1021/la8023698.
21. Garner, J.; Inglis, S. R.; Hook, J.; Separovic, F.; Harding, M. M. A Solid-State NMR Study of the Interaction of Fish Antifreeze Proteins with Phospholipid Membranes. Eur Biophys J 2008, 37 (6), 1031–1038. https://doi.org/10.1007/s00249-008-0339-3.
22. Clarke, J. A.; Heron, A. J.; Seddon, J. M.; Law, R. V. The Diversity of the Liquid Ordered (Lo) Phase of Phosphatidylcholine/Cholesterol Membranes: A Variable Temperature Multinuclear Solid-State NMR and X-Ray Diffraction Study. Biophysical Journal 2006, 90 (7), 2383–2393. https://doi.org/10.1529/biophysj.104.056499.
23. Dlouhý, O.; Kurasová, I.; Karlický, V.; Javornik, U.; Šket, P.; Petrova, N. Z.; Krumova, S. B.; Plavec, J.; Ughy, B.; Špunda, V.; Garab, G. Modulation of Non-Bilayer Lipid Phases and the Structure and Functions of Thylakoid Membranes: Effects on the Water-Soluble Enzyme Violaxanthin de-Epoxidase. Sci Rep 2020, 10 (1), 11959. https://doi.org/10.1038/s41598-020-68854-x.
Figures