1534
Measuring Quadrupolar Splittings in Human Lower Leg and Forearm Muscle after Deuterium Oxide Loading1School of Medicine, University of Nottingham, Nottingham, United Kingdom, 2NIHR Nottingham Biomedical Research Centre, University of Nottingham, Nottingham, United Kingdom, 3Sir Peter Mansfield Imaging Centre, University of Nottingham, Nottingham, United Kingdom, 4School of Physics, University of Nottingham, Nottingham, United Kingdom, 5School of Life Sciences, University of Nottingham, Nottingham, United Kingdom, 6MRC-Versus Arthritis Centre for Musculoskeletal Ageing Research, University of Nottingham, Nottingham, United Kingdom
Synopsis
Keywords: Muscle, Deuterium, Quadrupolar effects
Motivation: Although it is known that deuterium (2H) quadrupolar splittings arise in muscle, it is not clear to what extent they can be used to characterise tissues.
Goal(s): To investigate quadrupolar splittings by measuring angular dependence and double-quantum-filtered signals, and to determine whether splittings were related to specific muscles.
Approach: Deuterium oxide loading was used to increase the 2H signals. The angular dependence of the splitting was measured in the forearm and DQF spectra were acquired on forearm and lower leg muscles.
Results: Evidence was obtained for quadrupolar splittings which were shown to depend on angle and muscle group, and exhibited DQF spectra.
Impact: Deuterium quadrupolar splittings have the potential to characterise muscle fibres in vivo. Understanding the origin of the splittings, and factors affecting their magnitude, could lead to new or complementary methods in musculoskeletal or physiological investigations.
Background
Partially ordered biological tissues can be studied via deuterium (2H) magnetic resonance of water because the quadrupolar interaction produces a frequency doublet which is sensitive to the effect of ordering on the time-averaged direction of the local electric field gradient with respect to the B0-direction1,2. The low natural abundance of deuterium (0.015%) means that performing measurements in vivo can be challenging, even at high field3. Additionally, tissues typically contain multiple water compartments and spectra are complicated by the superposition of ordered (anisotropic) and disordered (isotropic) signal components. These issues were investigated by measuring the angular dependence of the 2H quadrupolar splittings from the lower leg and forearm muscles in healthy human participants who had ingested D2O to enrich their deuterium levels to ~100x natural abundance, and by comparing to measurements employing double-quantum filtering (DQF).Methods
Deuterium spectroscopy and chemical shift imaging (CSI) were performed on the lower leg and forearm in three human volunteers, using in-house-built 2H coils resonating at 19.6 MHz, interfaced to a Philips Achieva 3T scanner. A 16cm-diameter saddle coil was used for the lower leg, and a 15cm-diameter Helmholtz coil was used for the forearm.Deuterium 3D CSI images were acquired with resolution 10x10x10 mm3, TR/TE=500/6.2 ms, sampling bandwidth 750 Hz, 256 samples, 2 averages. Images were acquired in each subject with the lower leg parallel to the field and with the forearm at 10 different angles (~0 – 90°) to the field.
DQF deuterium spectra were acquired from a 2 cm axial slice of the lower leg or forearm, using hard pulses in combination with outer-volume saturation. Spectra were obtained via an anti-phase DQF sequence4 whereby the quadrupole-doublet peaks acquire a relative phase of 180°. DQF spectra were acquired for a range of values of the creation time, $$$1\leq\tau\leq36$$$ ms, with TR/TE=1000/0.58 ms, sampling bandwidth 3000 Hz, 1024 samples, 56 averages.
Deuterium 2D CSI data were also acquired from a single 2-cm axial slice, using outer-volume suppression. CSI data were acquired for the pulse-acquire sequence, and using the anti-phase DQF sequence (DQF-CSI) with $$$\tau=5$$$ ms. The in-plane resolution was 10x10 mm2, TR/TE=1000/2 ms, 256 samples, bandwidth 750 Hz, 4 averages (CSI), 8 averages (DQF-CSI).
Results
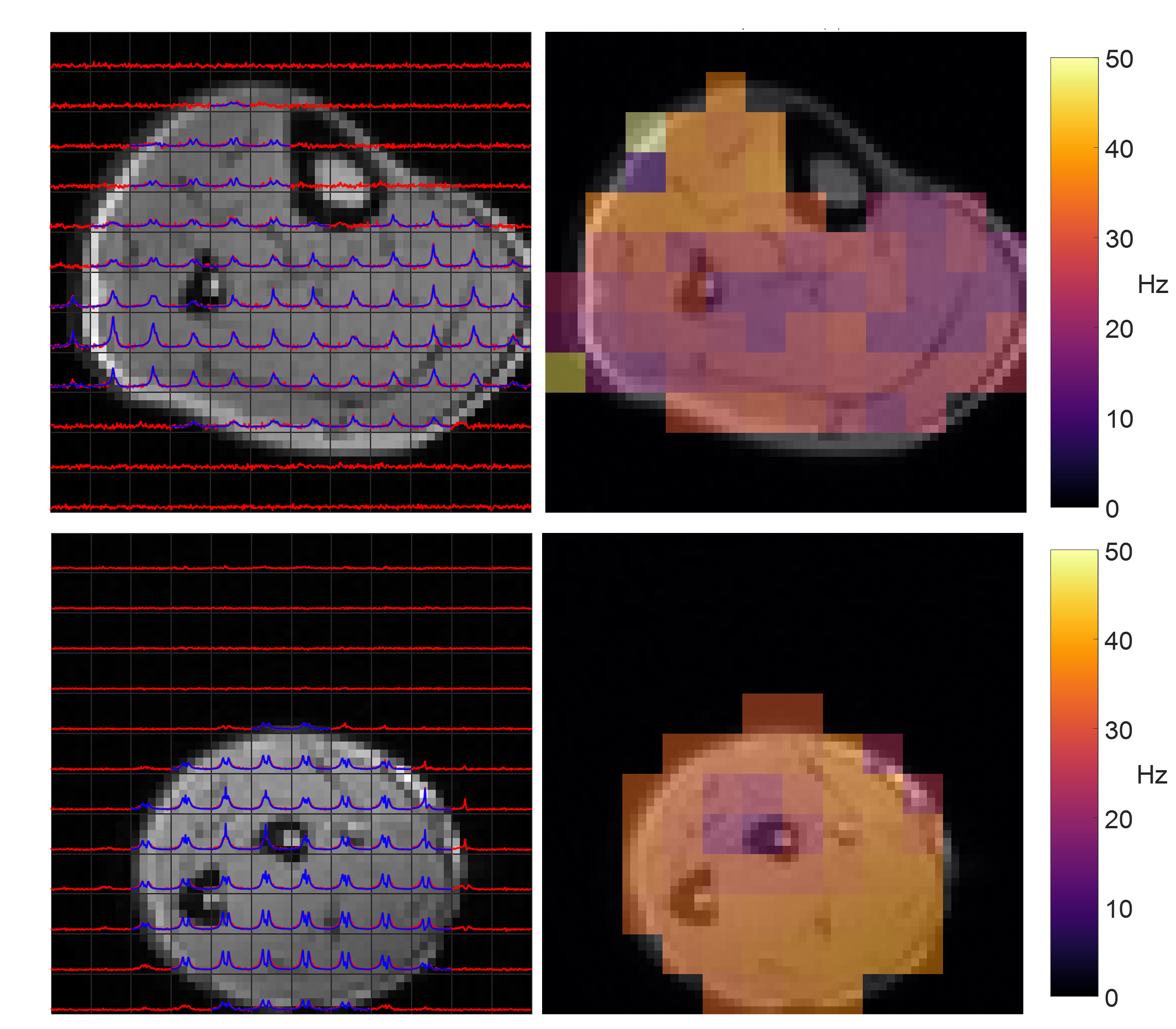
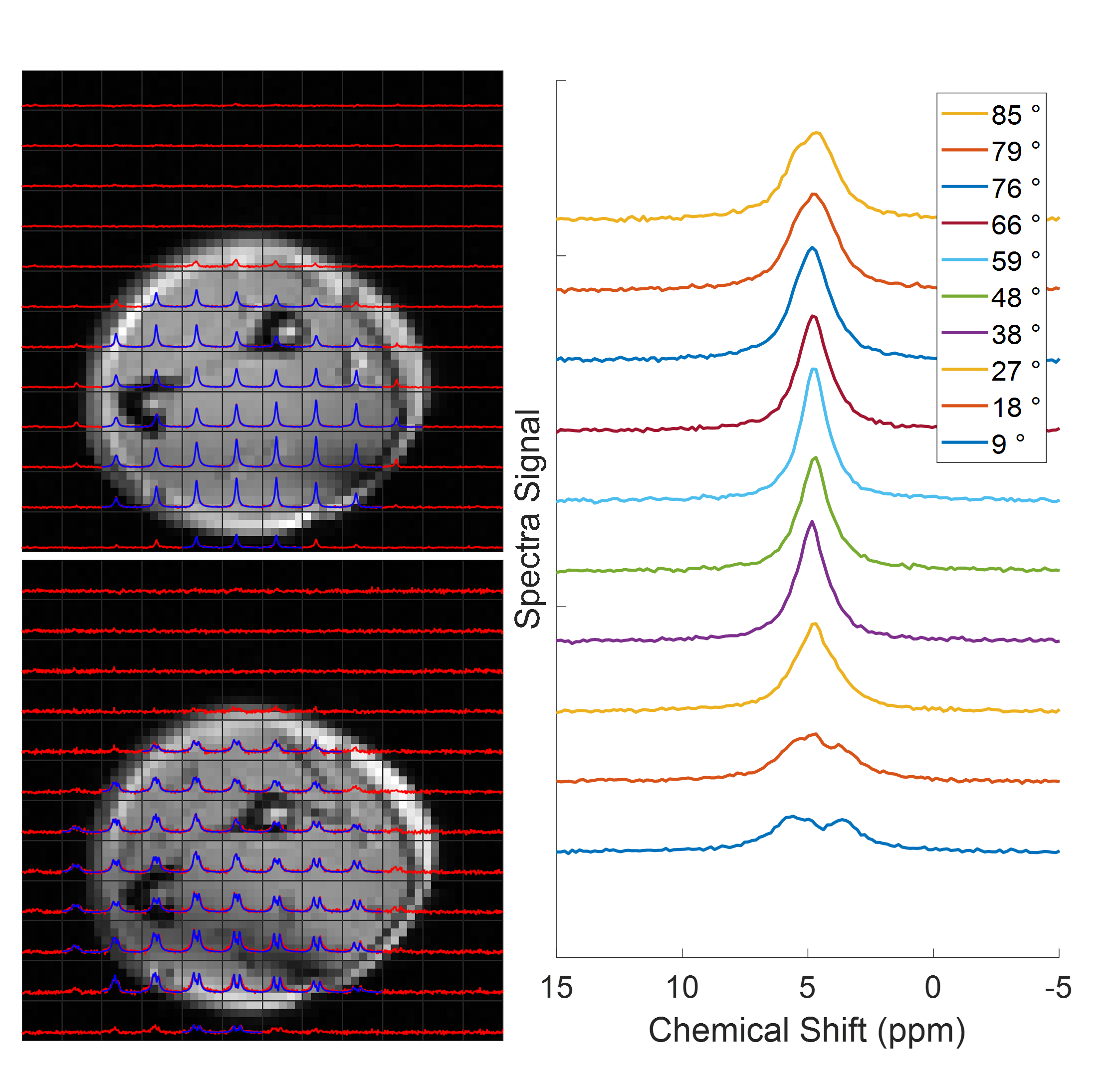
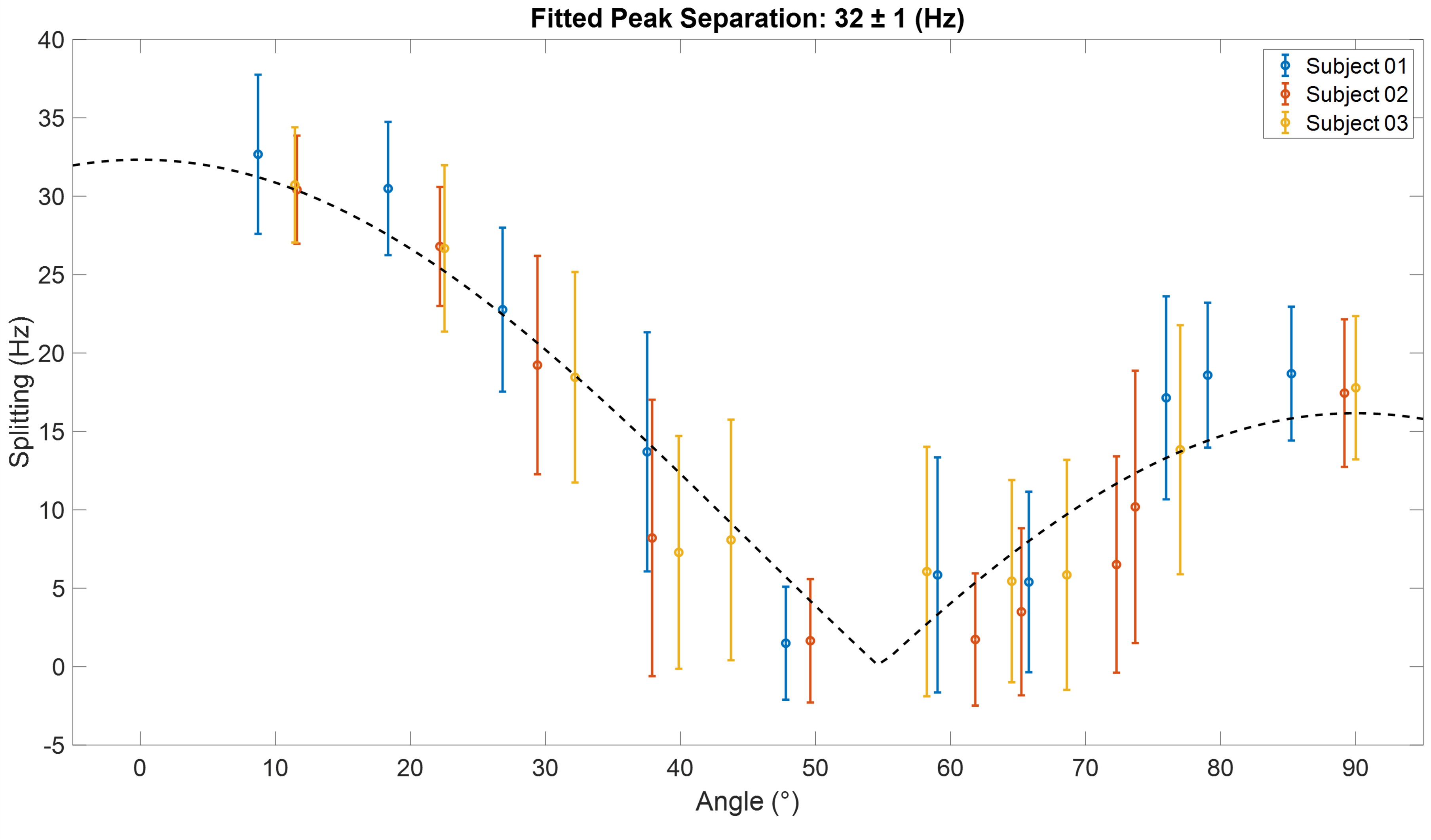
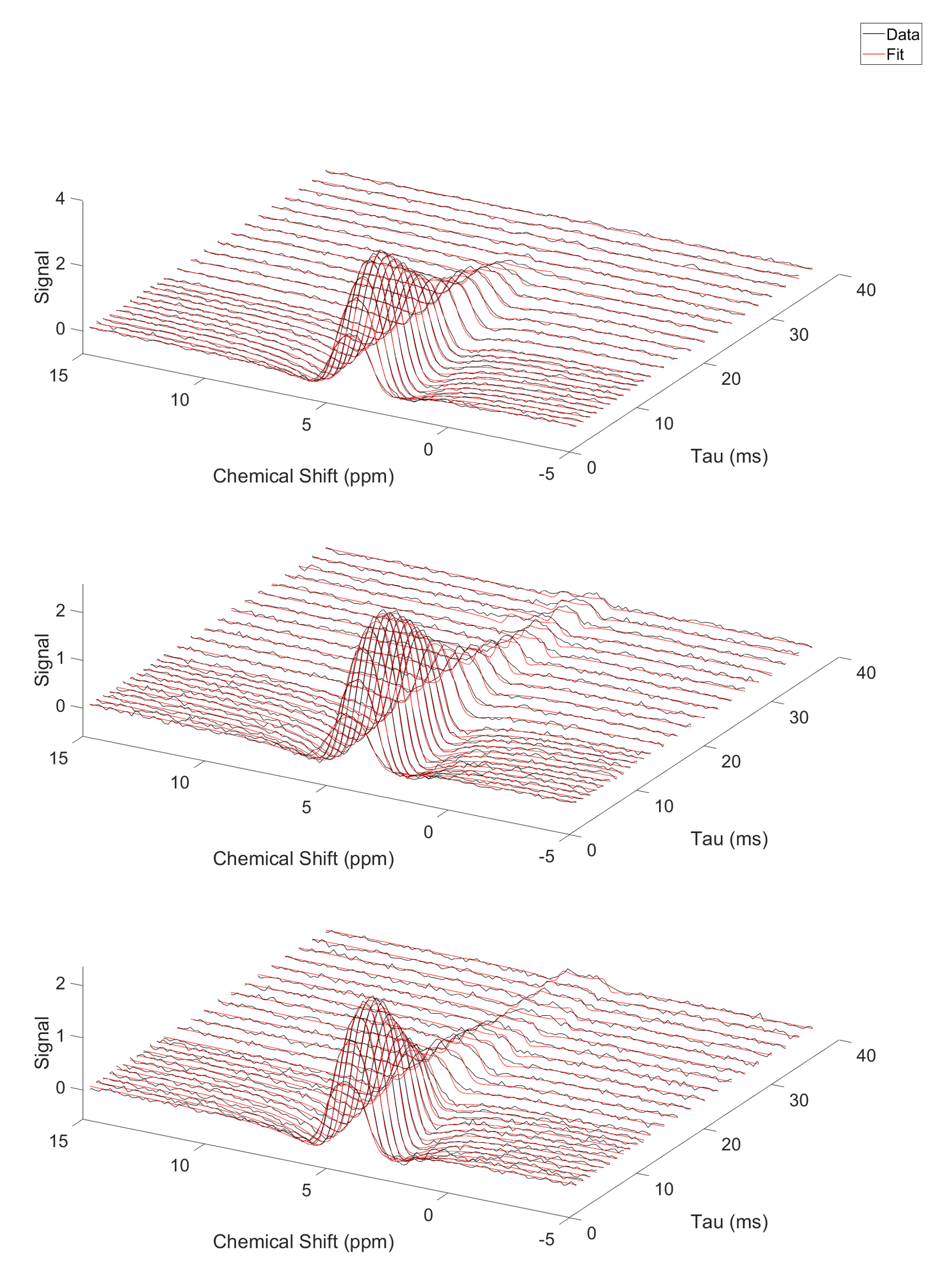
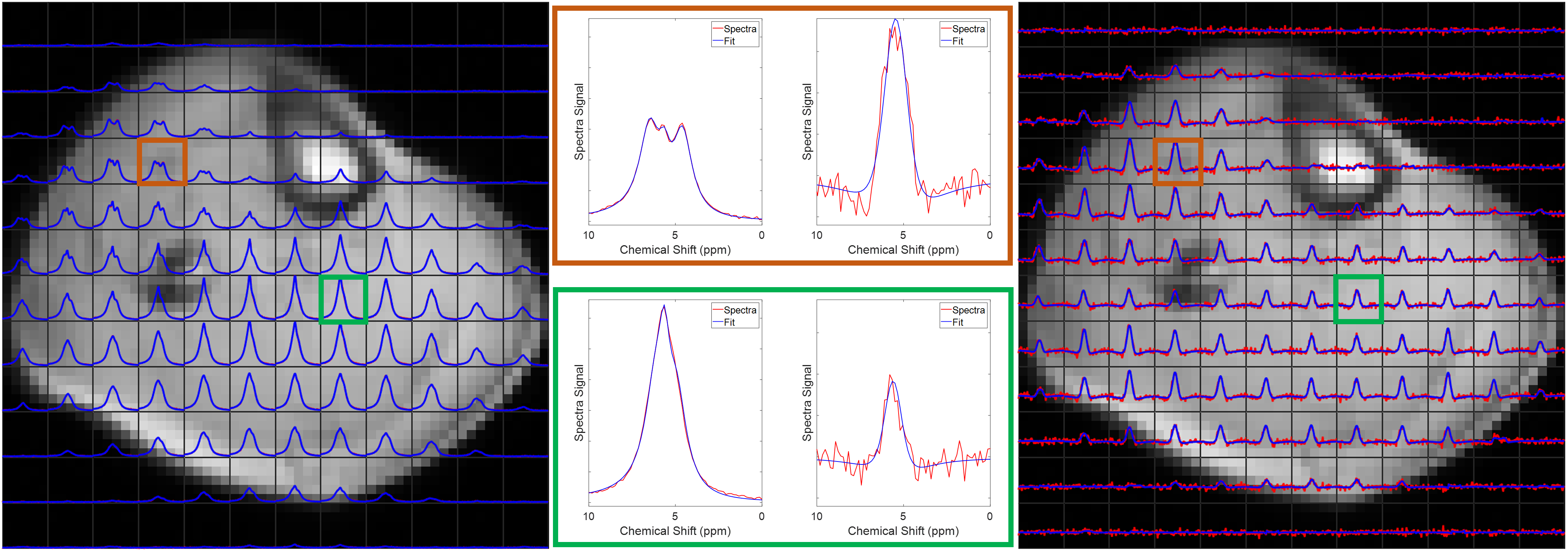
Figure 1 shows single slices from 3D CSI acquisitions of the lower leg and forearm, with the limb approximately aligned with the B0-direction. Doublets can be observed in many voxels, with residual quadrupolar splittings of 20 – 40 Hz. Figure 2 shows how the spectra from forearm muscles change in shape as the limb is oriented at different angles to B0, and Figure 3 plots the averaged quadrupolar splitting frequencies against angle. A fit to the expected variation as $$$f_q(3\cos^2(\theta)-1)/2$$$ provided an average value for all voxels and participants of $$$f_q=32\pm 1$$$ Hz.DQF spectra from the forearm are plotted as a function of creation time in Figure 4, and Figure 5 shows a comparison of 2D CSI and DQF-CSI data for the lower leg, highlighting spectra in individual voxels in the tibialis anterior and soleus.
Discussion
The residual quadrupolar splitting of the deuterated water spectrum is evidence of some local ordering of the tissue, and has been previously observed in muscle, tendon, cartilage, and nerves3—6. The CSI spectra in Figure 1 show that the splittings and lineshapes are spatially dependent in the lower leg, and the pattern of splittings roughly follows the different muscle groups, with the most prominent splittings occurring in the region of the tibialis anterior. This is partly because the fibres of this muscle align closely3 with the B0-direction, but it could also indicate a more ordered environment in which the water resides. As also seen in Figure 1, the muscles in the forearm appear to present more homogeneously. Evidence that the observed splittings show an angular dependence that is consistent with a quadrupolar interaction is provided in Figures 2 and 3. The measured value of $$$f_q$$$ is in agreement with previous 7T measurements in the lower leg3 and with values found from fitting to the DQF spectra (Fig. 4).The anti-phase DQF sequence produces spectra whose peaks have opposite phases. Consequently, signals with zero or very small splittings will vanish or be very small, and this provides a way of displaying only signals with non-zero splittings. These signals are shown in Figure 4, which as expected roughly follow the form $$$Ae^{-2\tau/T_2}\sin(2\pi f_q\tau)$$$. The utility of the DQF sequence is illustrated in Figure 5, where spectra with isotropic peaks in addition to a doublet (tibialis anterior) or no clear doublet (soleus) provide non-vanishing DQF spectra of varying amplitude.
Acknowledgements
This research was funded by the NIHR Nottingham Biomedical Research Centre. DJC’s PhD studies are funded by the Precision Imaging Beacon at the University of Nottingham.References
1. Seelig J. Deuterium magnetic resonance: theory and application to lipid membranes. Quarterly Reviews of Biophysics. 1977; 10(3): 353 – 418.
2. Eliav U, Navon G. Multiple Quantum MRS. eMagRes. 2016; 5(1). DOI: 10.1002/9780470034590.emrstm1448
3. Gursan A, Froeling M, Hendriks AD, Welting D, Kentgens APM, Klomp DWJ, Prompers JJ. Residual quadrupolar couplings observed in 7 Tesla deuterium MR spectra of skeletal muscle. Magnetic Resonance in Medicine. 2021; 00: 1 – 9. DOI: 10.1002/mrm.29053
4. Sharf Y, Eliav U, Shinar H, Navon G. Detection of anisotropy in cartilage using 2H double-quantum-filtered NMR spectroscopy. Journal of Magnetic Resonance. 1995; B107: 60 – 67.
5. Perea W, Cannella M, Yang J, Vega, AJ, Polenova T, Marcolongo M. 2H double quantum filtered (DQF) NMR spectroscopy of the nucleus pulposus tissues of the intervertebral disc. Magnetic Resonance in Medicine. 2007; 57: 990 – 999.
6. Eliav U, Wehli FW, Navon G. New insight into the organisation of myelin water using deuterium NMR. Magnetic Resonance in Medicine. 2020; 84(2): 535 – 541.
Figures