1361
Two-axon population (TAP) modelling for large axon diffusion imaging in the peripheral nervous system1NMR Research Unit, Queen Square MS Centre, Department of Neuroinflammation, UCL Queen Square Institute of Neurology, Faculty of Brain Sciences, University College London, London, United Kingdom, 2Radiomics Group, Vall d’Hebron Institute of Oncology, Vall d’Hebron Barcelona Hospital Campus, Barcelona, Spain, 3Department of Brain and Behavioural Sciences, University of Pavia, Pavia, Italy, 4Digital Neuroscience Centre, IRCCS Mondino Foundation, Pavia, Italy
Synopsis
Keywords: Diffusion Modeling, Microstructure, Sciatic nerve, axon radius, Multiple Sclerosis
Motivation: The stick (zero-radius cylinder), a standard diffusion MRI model for the intra-axonal brain white matter signal, may not be suitable in the peripheral nervous system (PNS), due to co-existence of small and large axons.
Goal(s): To test the feasibility of replacing the stick model with a two-axon population (TAP) approach, accounting for large axons, in the PNS.
Approach: We compared TAP versus standard stick modelling in simulations, and derived TAP metrics in the sciatic nerve in vivo.
Results: TAP enables more accurate parameter fitting than stick modelling, and is shown to be feasible in vivo in healthy controls and multiple sclerosis patients.
Impact: We propose two-axon population (TAP) diffusion-weighted MRI, a method tailored for imaging of the peripheral nervous system, characterised by co-existence of small and large axons. TAP may enable more accurate microstructural imaging than standard approaches, providing non-invasive markers of neurophysiology.
Introduction
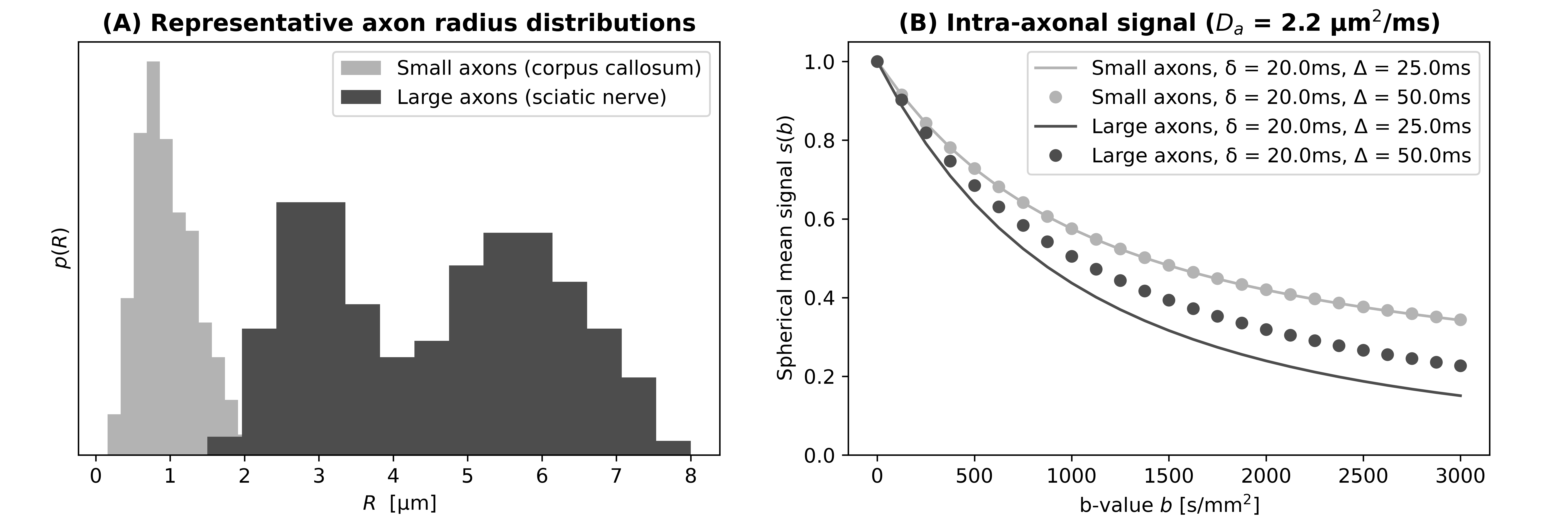
The stick (zero-radius cylinder) is a common diffusion-weighted (DW) signal model used for axons1-5. The model is suitable for radii up to approximately 3 µm in clinical systems6, i.e., for most brain white matter. However, a mixture of small and large axons is found in the peripheral nervous system (PNS7; radii as large as 8 µm). For such large axons, the DW signal deviates from the idealised stick signal8, featuring stronger attenuation and diffusion-time dependence (Fig. 1). This may bias the estimation of key microstructural parameters such as the intra-axonal (IA) signal fraction $$$f_a$$$, a promising marker of axonal loss. To address this issue, we investigated the feasibility of replacing the stick model with a two-axon population (TAP) approach accounting for large axons. Specifically, we compared TAP and stick signal modelling in simulations, and tested the application of TAP in the PNS in vivo.Methods
SimulationsWe compared $$$f_a$$$ estimates of IA signal fraction obtained through both TAP and standard “stick” modelling in synthetic signals.
Signal synthesis We synthesised the total DW signal by summing IA and extra-axonal (EA) signal contributions ($$$s_a$$$ and $$$s_e$$$) as
$$s\,\,=\,\,f_a \,s_a\,\,+\,\,(1-f_a)\, s_e.\,\,\,\,\,(1)$$
$$$s_a$$$ integrates contributions from a distribution of axonal radii, i.e.,
$$s_a\,\,=\,\, c\int_{0}^{\infty}R^2\,h(R,D_a,b,\delta,\Delta)\,p(R;\eta,\zeta)\,dR.\,\,\,\,\,(2)$$
Above, $$$p(R;\eta,\zeta)$$$ is a gamma distribution8,9 (shape: $$$\eta$$$; scale: $$$\zeta$$$; mean: $$$R_m=\eta\,\zeta$$$; variance: $$$R_{s}^{2}=\eta\,\zeta^2$$$), and $$$c$$$ ensures $$$s_a(b=0)=1$$$. $$$h(R,D_a,b,\delta,\Delta)$$$ is the powder-averaged signal4,10 for diffusion within a cylindrical axon of radius $$$R$$$ and IA diffusivity $$$D_a$$$, i.e.,
$$h(R,D_a,b,\delta,\Delta)\,\,=\,\frac{\sqrt{\pi}}{2}\,e^{-b\,D_p(R,D_a,\delta,\Delta)}\,\frac{\mathrm{erf}\Big( \sqrt{b\big(D_a - D_p(R,D_a,\delta,\Delta)\big)} \Big)}{\sqrt{b\big(D_a - D_p(R,D_a,\delta,\Delta)\big)}}.\,\,\,\,\,(3)$$
Above,
$$D_p(R,D_a,\delta,\Delta)\,\,=\,\,\frac{2}{D_a\delta^2(\Delta-\delta/3)}\,\sum_{m=1}^{\infty}\,\frac{\big(2D_a\alpha_m^2\delta\,-2\,+2e^{-D_a\alpha_m^2\delta}\,+2e^{-D_a\alpha_m^2\Delta}\,-e^{-D_a\alpha_m^2(\Delta-\delta)}\,-e^{-D_a\alpha_m^2(\Delta+\delta)}\big)}{\alpha_m^6(R^2\alpha_m^2-1)}\,\,\,\,\,(4)$$
is the IA perpendicular apparent diffusion coefficient (ADC)11, and $$$\alpha_m$$$ indicates the roots of $$$J_{1}^{'}(\alpha_m R) = 0$$$. Finally, the EA signal is generated as
$$s_e\,\,=\,\,e^{-b\,D_e},\,\,\,\,\,(5)$$
where $$$D_e$$$ is the EA ADC.
We synthesised signals for 1500 uniformly-distributed combinations of $$$f_a$$$, $$$D_a$$$, $$$D_e$$$, $$$R_m$$$, $$$R_s$$$ ([0; 1], [1.0; 3.0] µm2/ms, [0.5; 3.0] µm2/ms, [1; 6] µm, [0.3$$$R_m$$$; 0.7$$$R_m$$$]). The protocol was b = {0, 500, 1000, 1500, 2000, 2500, 3000} s/mm2 × Δ = {30, 50, 70} ms (fixing δ = 20ms). Signals were corrupted with Rician noise (b=0 signal-to-noise-ratio: 20).
$$$f_a$$$ estimation We fitted an IA/EA model
$$ s\,\,=\,\,f_a\,s_a\,\,+\,\,(1-f_a)\,e^{-b\,D_e},\,\,\,\,\,(6)$$
comparing two implementations of $$$s_a$$$:
- TAP: $$$s_a$$$ is written as
$$s_a\,\,=\,\,f_l\,h(R_l,D_a,b,\delta,\Delta)\,\,+\,\,(1-f_l)\,h(0,D_a,b,\delta,\Delta),\,\,\,\,\,(7)$$
to include a mixture of "sticks" and large axons, with signal fraction $$$f_l$$$ and characteristic radius $$$R_l$$$. Note that in the “stick” model $$$h(0,D_a,b,\delta,\Delta)$$$ the IA perpendicular ADC vanishes, since in Eq. 4 above, $$$D_p(R,D_a,\delta,\Delta) \rightarrow 0$$$ for $$$R \rightarrow 0$$$.
- Stick: $$$s_a$$$ follows the standard zero-radius model
$$s_a\,\,=\,\,h(0,D_a,b,\delta,\Delta).\,\,\,\,\,(8)$$
Estimated/reference $$$f_a$$$ were compared through bias and dispersion (variability) indices.
In vivo demonstration
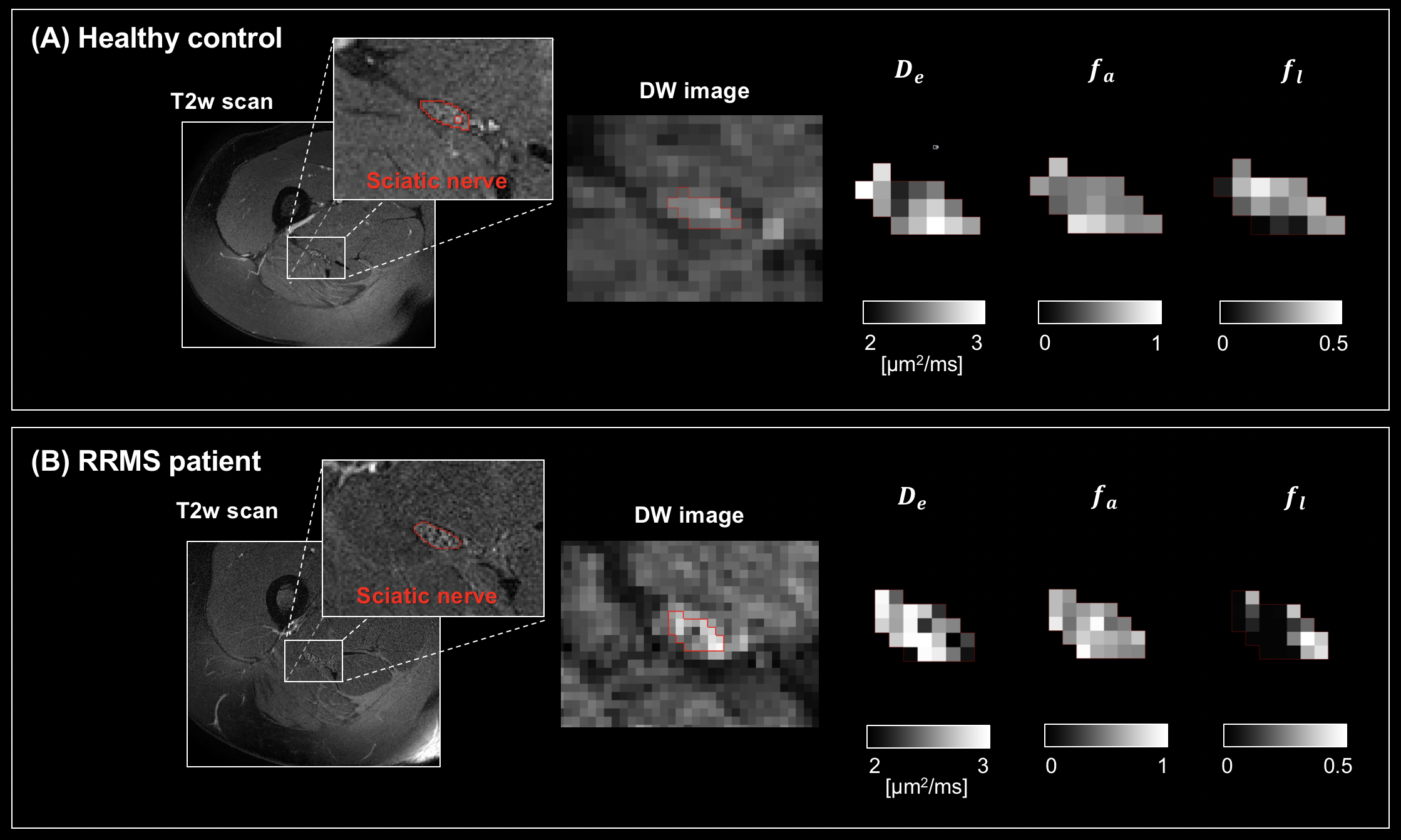
We investigated TAP modelling in vivo by analysing retrospectively sciatic nerve scans from 24 healthy volunteers (mean age 36.4 years, range 26-53, 17 female) and 23 people with relapsing-remitting multiple sclerosis (RRMS; mean age 40.0 years, range 30-59, 13 female). DW images, acquired at 3T (Philips Ingenia CX, b = {0, 700, 1200, 2000} s/mm2, δ = 15.06 ms, Δ = 37.55 ms, resolution 1×1×10 mm3, TE=80 ms; TR=4000 ms), were pre-processed12,13, and $$$D_e$$$, $$$f_a$$$ and $$$f_l$$$ computed. Mean values within the manually-segmented sciatic nerve were compared between groups via linear regression, adjusting for age, gender, and body mass index.
Results and discussion
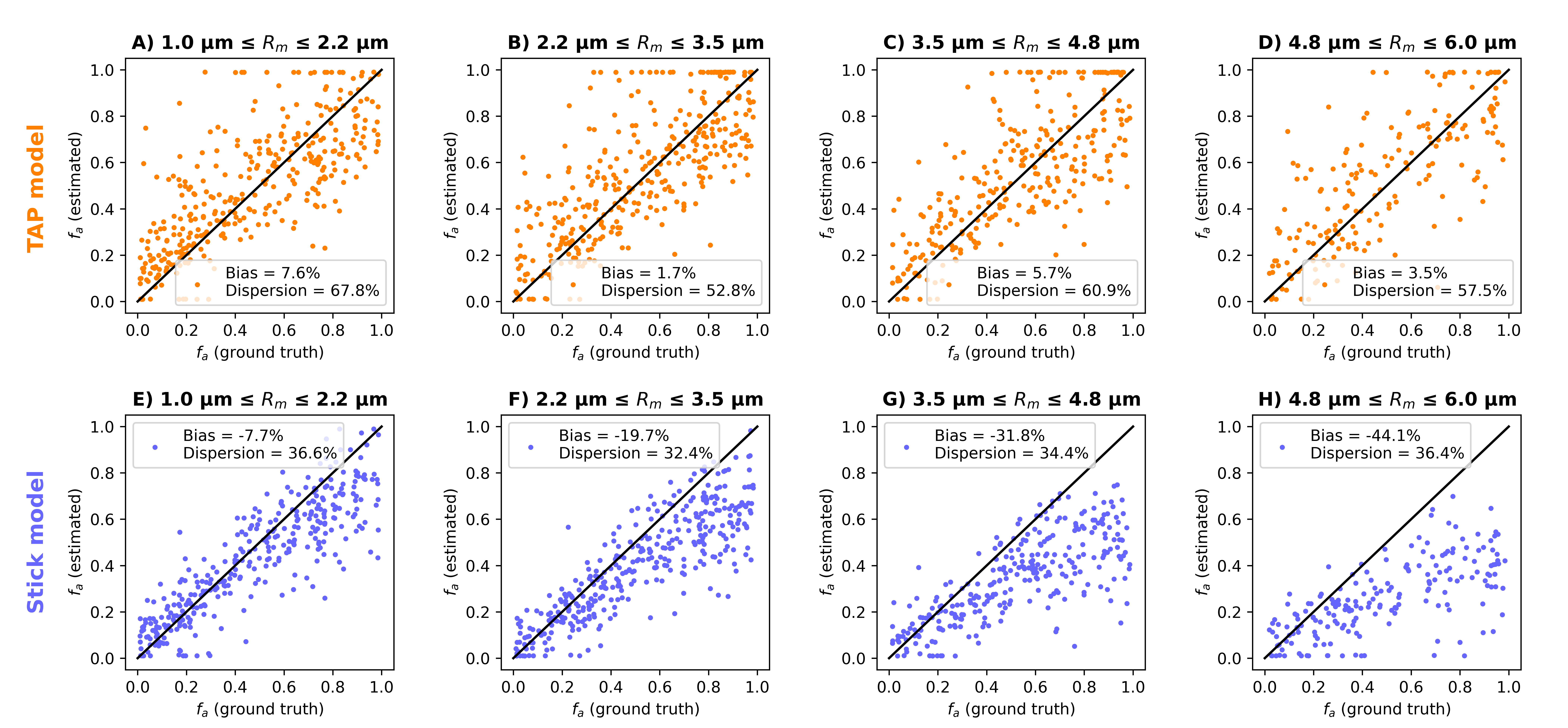
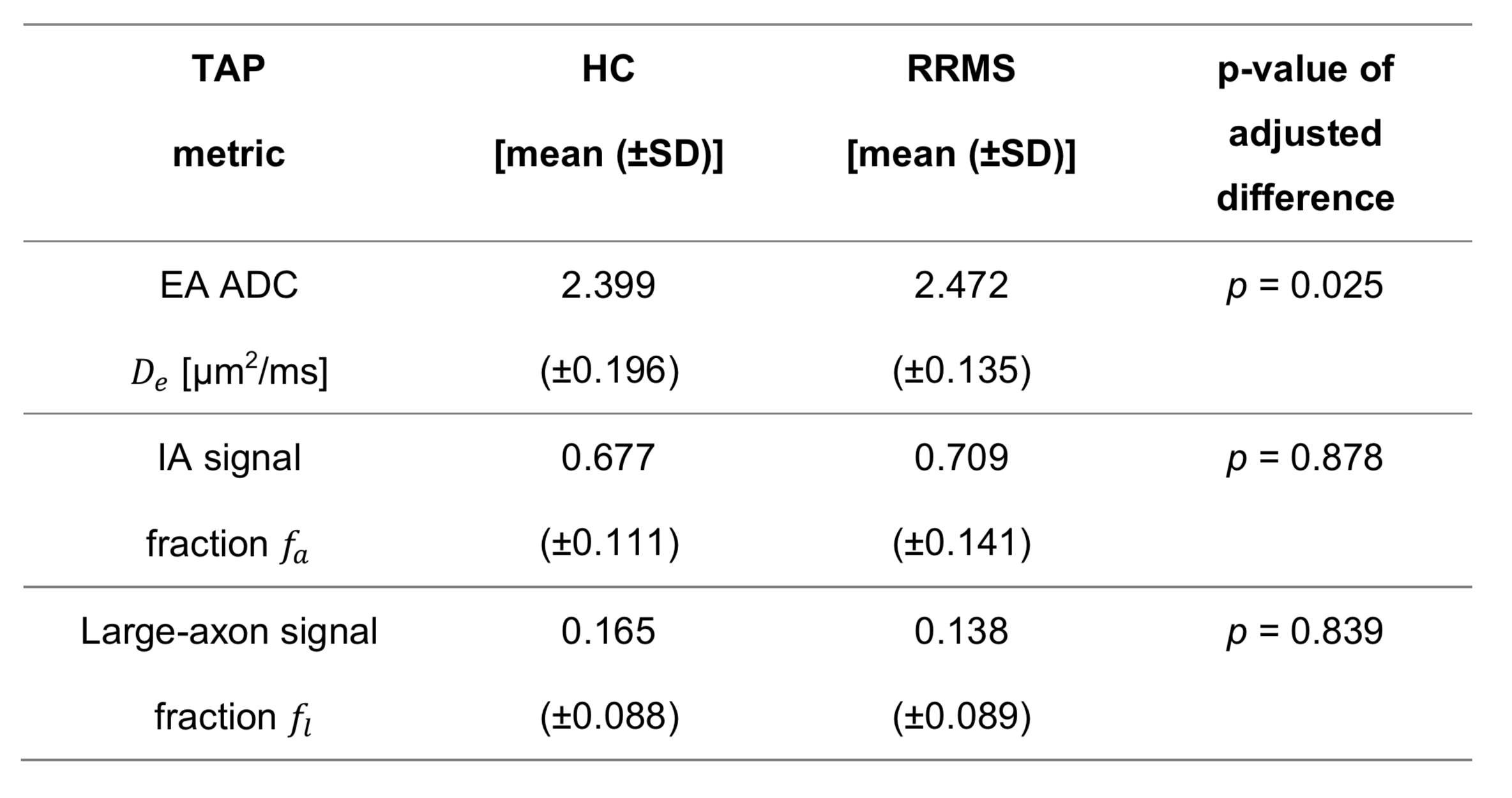
Fig. 2 shows scatter plots visualising $$$f_a$$$ predictions for axons with increasingly large mean radius $$$R_m$$$. For the smallest axons, both TAP and the stick models provide accurate estimates of $$$f_a$$$. However, as $$$R_m$$$ increases, stick estimates become increasingly biased, underestimating ground truth $$$f_a$$$, unlike TAP. This result points towards the need for accounting for finite axon radius in the presence of large axons. Finally, TAP $$$f_a$$$ shows higher variability than stick estimates, owing to the increased number of unknown tissue parameters, which makes TAP more difficult to fit than the stick model.Fig. 3 shows examples of TAP maps $$$D_e$$$, $$$f_a$$$ and $$$f_l$$$ in vivo in a representative control and a representative RRMS patient, while Table 1 reports mean/standard deviation values in the two groups. In both patients and controls, voxels with high $$$f_l$$$ values, consistent with the presence of a significant number of large axons, are seen. Statistically significant patient-control differences are observed for the EA ADC $$$D_e$$$, with higher $$$D_e$$$ in patients than in controls (p = 0.025), compatible with early demyelination. This finding suggests that TAP metrics may be useful biomarkers for non-invasive neurophysiological assessment of PNS microstructure, as in MS applications.
Conclusions
TAP modelling can reduce biases in IA signal fraction estimation in the presence of large axons. It provides microstructural metrics that may become useful biomarkers in PNS applications, as in the characterisation of preferential large/small axon degeneration.Acknowledgements
FG and RB are joint first authors with equal contribution. This work is financially supported by the National Brain Appeal’s Innovation fund (https://www.nationalbrainappeal.org). FG receives the support of a fellowship from ”la Caixa” Foundation (ID 100010434). The fellowship code is “LCF/BQ/PR22/11920010”. RB receives a scholarship from Chiang Mai University, Chiang Mai, Thailand. CGWK receives funding from Horizon 2020 (Research and Innovation Action Grants Human Brain Project 945539 (SGA3)), BRC (#BRC704/CAP/CGW), MRC (#MR/S026088/1), Ataxia UK, Rosetrees Trust (#PGL22/100041 and #PGL21/10079). CGWK is a shareholder in Queen Square Analytics Ltd.References
1. Behrens TEJ, et al. Characterization and propagation of uncertainty in diffusion-weighted MR imaging. Magnetic Resonance in Medicine 2003, 50(5): 1077-1088.
2. Zhang H, et al. NODDI: practical in vivo neurite orientation dispersion and density imaging of the human brain. Neuroimage 2012, 61(4): 1000-1016.
3. Novikov DS, et al. Rotationally-invariant mapping of scalar and orientational metrics of neuronal microstructure with diffusion MRI. NeuroImage 2018, 174: 518-538.
4. Palombo M, et al. SANDI: a compartment-based model for non-invasive apparent soma and neurite imaging by diffusion MRI. Neuroimage 2020, 215: 116835.
5. Jelescu IO, et al. Neurite Exchange Imaging (NEXI): A minimal model of diffusion in gray matter with inter-compartment water exchange. NeuroImage 2022, 256: 119277.
6. Nilsson M, et al. Resolution limit of cylinder diameter estimation by diffusion MRI: The impact of gradient waveform and orientation dispersion. NMR in Biomedicine 2017, 30(7): e3711.
7. Romero E, et al. Automatic morphometry of nerve histological sections. Journal of Neuroscience Methods 2000, 97(2): 111-122.
8. Grussu F, et al. Relevance of time‐dependence for clinically viable diffusion imaging of the spinal cord. Magnetic Resonance in Medicine 2019, 81(2): 1247-1264.
9. Assaf Y, et al. AxCaliber: a method for measuring axon diameter distribution from diffusion MRI. Magnetic Resonance in Medicine 2008, 59(6): 1347-1354.
10. Kaden E, et al. Quantitative mapping of the per‐axon diffusion coefficients in brain white matter. Magnetic Resonance in Medicine 2016, 75(4): 1752-1763.
11. Vangelderen P, et al. Evaluation of restricted diffusion in cylinders. Phosphocreatine in rabbit leg muscle. Journal of Magnetic Resonance Series B 1994, 103(3): 255-260.
12. Veraart J, et al. Denoising of diffusion MRI using random matrix theory. Neuroimage 2016, 142: 394-406.
13. Andersson JL, et al. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. Neuroimage 2016, 125: 1063-1078.
Figures