1313
Lifespan Iron Accumulation in Dopaminergic Neurons Studied by Quantitative MRI and X-ray Fluorescence1Neurophysics Department, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany, 2Paul Flechsig Institute of Neuropathology and Brain Research, Medical Faculty University Leipzig, Leipzig, Germany, 3Deutsches Elektronen-Synchrotron DESY, Hamburg, Germany, 4Full brain picture Analytics, Leiden, Netherlands, 5Ape Social Mind Lab, Institute of Cognitive Science Marc Jeannerod, Lyon, France, 6Taï Chimpanzee Project, Centre Suisse de Recherches Scientifiques, Abidjan, Cote D'ivoire, 7Felix Bloch Institute for Solid State Physics, Faculty of Physics and Earth System Sciences, Leipzig University, Leipzig, Germany, 8Wellcome Centre for Human Neuroimaging, Institute of Neurology, University College London, London, United Kingdom
Synopsis
Keywords: Microstructure, Quantitative Imaging, Iron, Substantia nigra, R2*
Motivation: Dopaminergic neurons require iron for their function but suffer from iron overload in age.
Goal(s): To non-invasively monitor the age-related iron accumulation in dopaminergic neurons, we investigated mechanisms of iron-induced MR contrast in the substantia nigra across the lifespan.
Approach: We combined quantitative MRI, X-ray fluorescence imaging and biophysical modelling in a unique animal model: ethically collected postmortem chimpanzee brains.
Results: The iron load of dopaminergic neurons and the effective transverse relaxation rates in the substantia nigra increased with age. The biophysical model accurately links the relaxation rate to the iron load and neuronal density, which demonstrated its suitability for ages above puberty.
Impact: Monitoring cell-specific iron concentrations of dopaminergic neurons and neuronal densities in the substantia nigra throughout the lifespan holds potential of an early neuroimaging biomarker for Parkinson’s disease.
Introduction
Dopaminergic neurons (DN) in the substantia nigra (SN) require sufficient levels of cellular iron for the synthesis of the neurotransmitter dopamine. They may suffer from iron overload in age1. In healthy DN, neuromelanin (NM) is the main iron storage and has a neuroprotective function: it chelates labile iron to prevent oxidative stress2. However, as iron accumulates in NM throughout life, iron-overloaded NM may release iron and become neurotoxic, posing a risk factor for Parkinson‘s disease (PD)1. Non-invasive methods for monitoring DN integrity and iron load are urgently needed for early-stage PD diagnosis and the development of curative treatments.Empirical models have related the transverse (R2) and effective transverse (R2*) relaxation rate to the total tissue iron concentration, opening the possibility for in-vivo monitoring of iron. These models, however, do not distinguish between ferritin-bound iron located in glial cells and NM-bound iron in DN3,4 and are therefore cannot disentangle iron accumulation related to different physiological and pathological processes. Recently, we demonstrated that iron in DN in the human SN pars compacta can be quantified with the help of a generative model linking R2 and R2* to DN density and cellular iron load5. However, this model has only been validated in postmortem tissue from elderly donors and the mechanisms of iron-driven contrast in MRI in younger individuals have not yet been explored. The main challenge is that postmortem brain samples from young human donors are rare and commonly used animal models are not useful due to short lifespans and absence of NM in rodent brains.
Herein, we characterized the iron accumulation in DN during the lifespan using a unique animal model: ethically collected postmortem brains of chimpanzees6. Using ultra-high resolution quantitative R2* maps and iron quantification by X-ray fluorescence (XRF), we extended the previously proposed biophysical model from the adult age to the entire hominoid lifespan5.
Methods
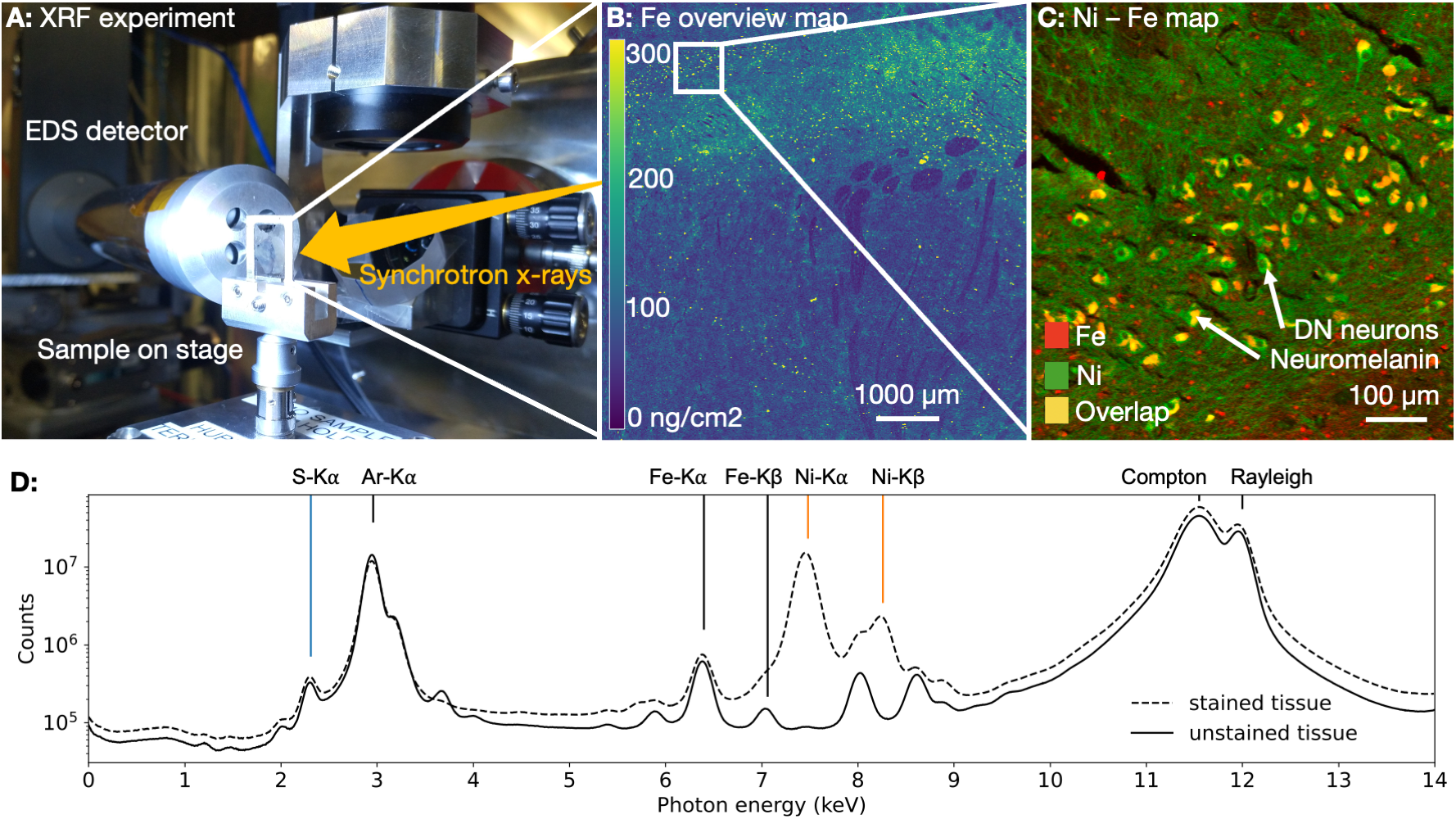
22 postmortem brains of chimpanzees (0.1-56years, 8 females) were collected within the Evolution of Brain Connectivity (EBC) project*6. Quantitative MRI (qMRI) was acquired on a 7T Terra MRI scanner (Siemens Healthineers). Automated parcellation of SN was achieved by Multi-contrast Anatomical Subcortical Structure Parcellation (MASSP7) adapted to non-human primate brains. Median R2* values in SN were used as a proxy for averaged iron concentrations in SN and were compared to literature values from humans3,7.For a subset of five brains (0.1;1.7;16;30;44years, 1 female), cellular iron quantification was performed on 10µm thick paraffin sections using XRF microscopy at PETRA III storage ring at the Deutsches Elektronen-Synchrotron (DESY). The biophysical model of R2*-contrast induced by DN based on the static dephasing theory was informed by cellular iron concentration and used to estimate the contribution of DN to R2* in the SN at different ages5,8.
Results
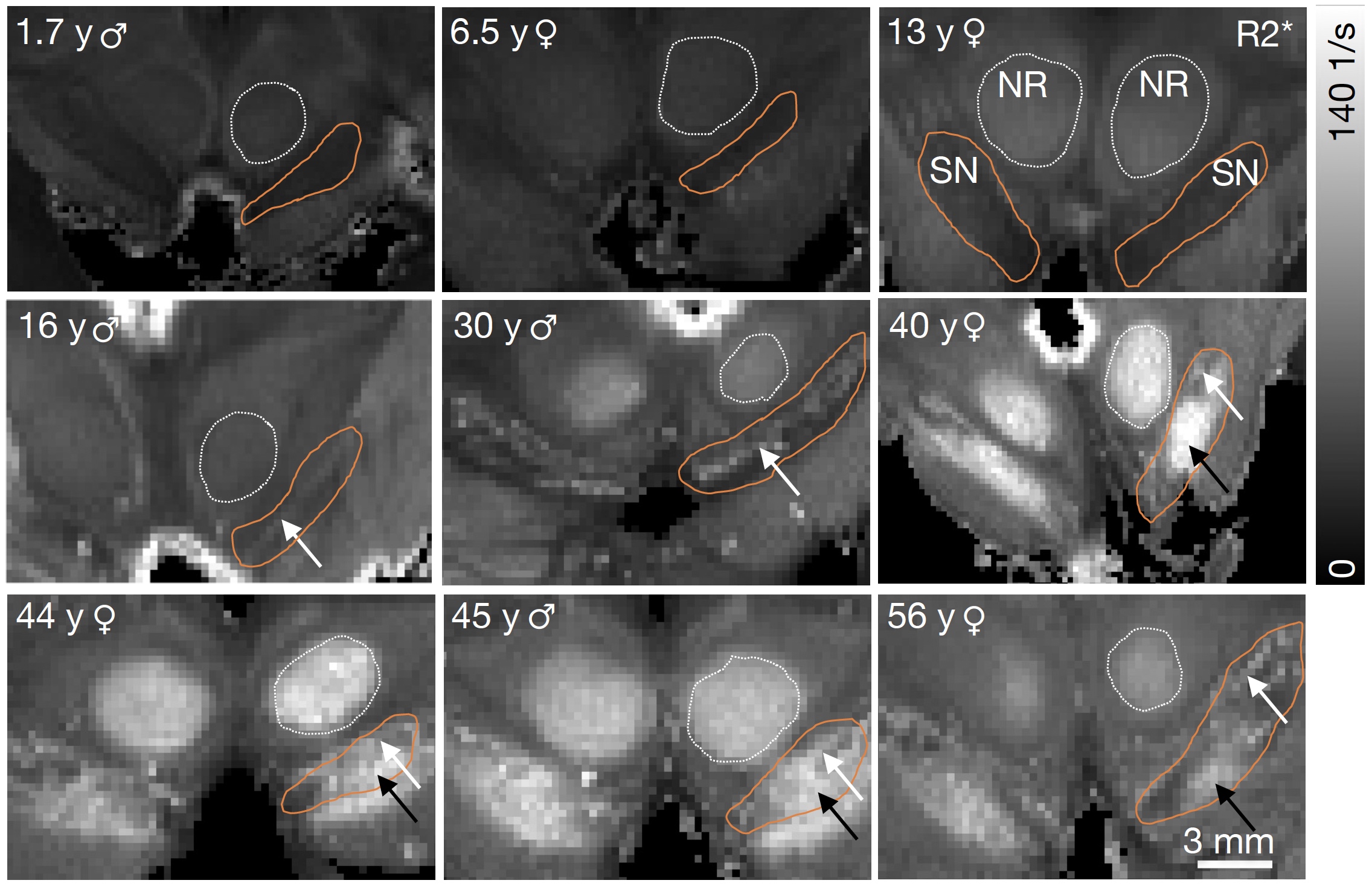
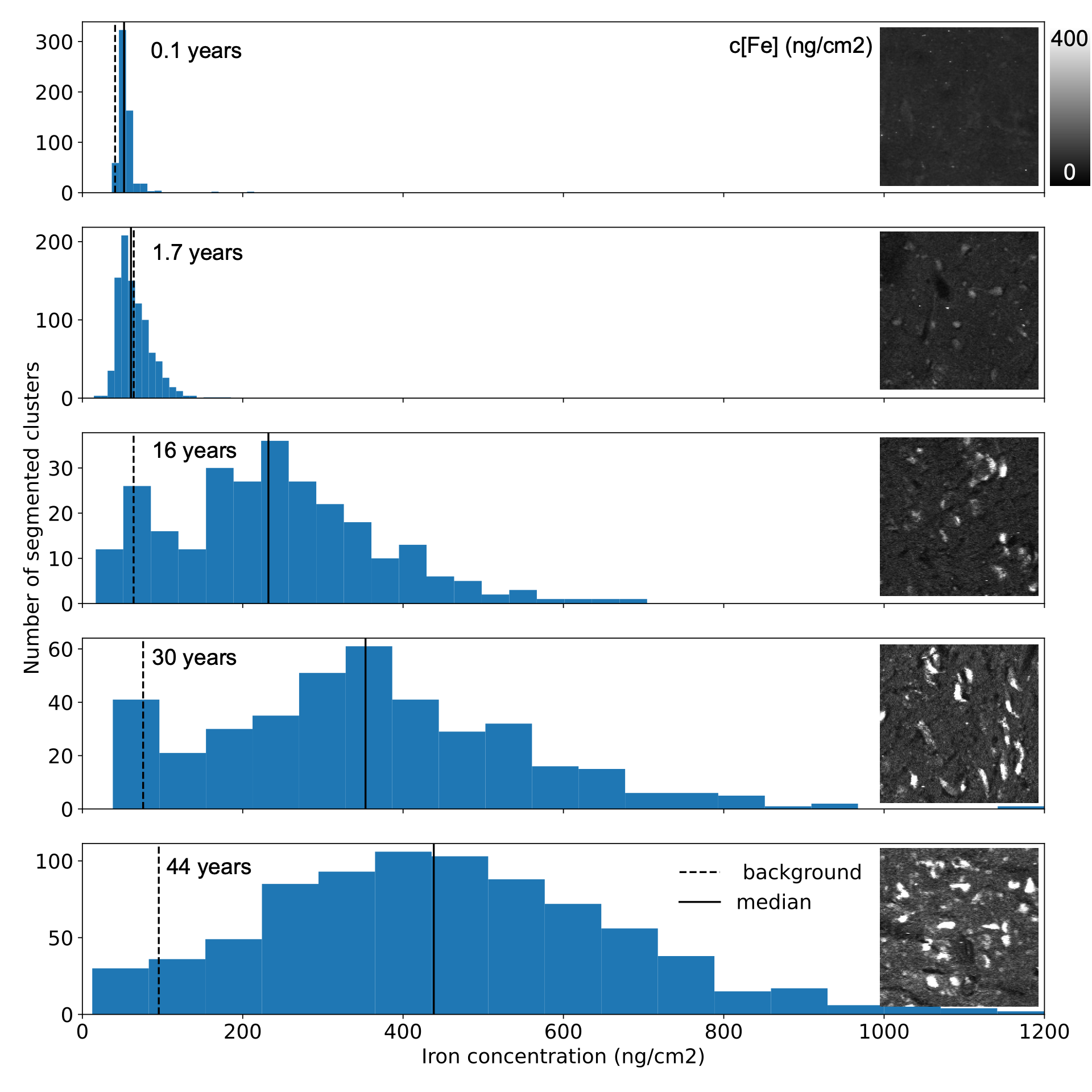
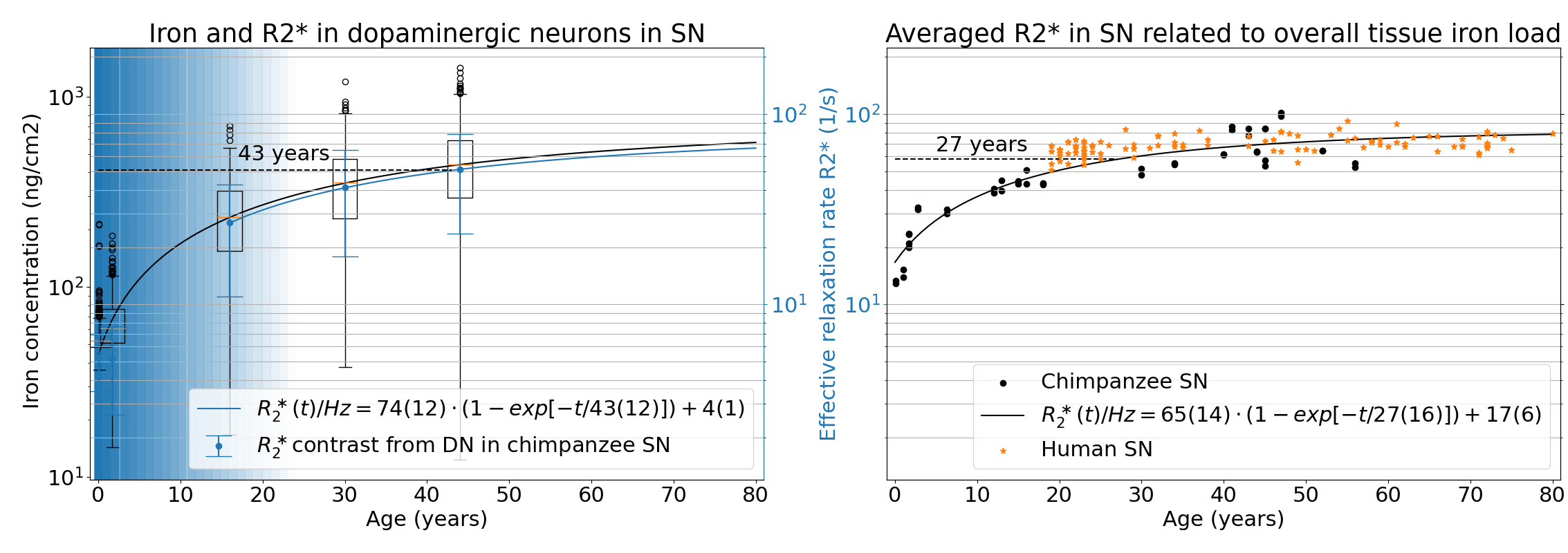
The NM accumulation in DN of chimpanzee brains across the lifespan was found to be qualitatively similar to literature reports from humans9 (Fig.1). Averaged R2*-rates in the SN, reflecting the averaged tissue iron concentration, increased with age. In adulthood, those rates were similar to values reported in humans, when differences in relaxation rates between postmortem and in-vivo tissue are taken into account5 (Figs.2,5). The similarity in NM accumulation and total iron levels in SN between the two species qualifies the chimpanzee as a suitable animal model to study the lifespan trajectory of human DN.In the subset of five animals, the iron concentration in NM clusters increased from 50(14)µg/g at birth to 440(220)µg/g at age (median(standard deviation), Figs.3,4). It was characterized by an exponential saturation with a time constant of 47(8)years (Fig.5). The static dephasing approximation accurately described the NM-bound-iron-induced contribution to R2* for adult chimpanzees aged 12-16years and older (Fig.5). The averaged tissue iron concentration in SN, estimated using averaged R2* values, increased with a time constant of 27(16)years, thus substantially faster than DN iron.
Discussion
For the first time, we quantified the age-related iron accumulation of NM-clusters in DN in the SN of our closest relatives, chimpanzees, using XRF. We linked them to qMRI parameters and compared them to the overall tissue iron accumulation. The different time courses for the accumulation of total tissue iron and NM iron in DN indicate different underlying physiological processes.The concordance between predicted microscopic R2* in DN and experimental macroscopic R2* in the entire SN underpins that the increase in R2* with age reflects the iron accumulation in the SN. We demonstrated that the biophysical model of iron-induced R2* in the SN is applicable after puberty opening the way for a non-invasive biomarker of DN density and iron load, which may be used to diagnose and monitor PD.
Acknowledgements
This project has received funding from the European Research Council (ERC n°616905), from the Federal Ministry of Education and Research (BMBF, 01ED2210), from the German Research Foundation (DFG, WE 5046/4-2, KI 1337/2-2) and the Max Planck Society. We acknowledge DESY (Hamburg, Germany), a member of the Helmholtz Association HGF, for the provision of experimental facilities. Parts of this research were carried out at PETRA-III and we would like to thank K. V. Falch and J. Garrevoet for assistance in using the microprobe at beamline P06. Beamtime was allocated for proposal I-20211534.
* EBC (Evolution of Brain Connectivity) consortium: Bala Amarasekaran (Tacugama Chimpanzee Sanctuary, Freetown, Sierra Leone) Caroline Asiimwe (Budongo Conservation Field Station, Uganda) Penelope Carlier (Tai Chimpanzee Project, CSRS, Abidjan, Cote d’Ivoire) Julian Chantrey (Veterinary Pathology and Preclinical Sciences, University of Liverpool, UK) Tobias Deschner (Institute for Cognitive Sciences, University of Osnabrueck, Germany; Ozouga, Loango Chimpanzee Project, Loango National Park, Gabon) Ariane, Düx (Robert Koch Institute, Berlin, Germany; Helmholtz Centre for Infection Research, Greifswald, Germany) Luke J. Edwards (Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany) Pawel Fedurek (School of Psychology, University of Stirling, UK; Budongo Conservation Field Station, Masindi, Uganda) Angela D. Friederici (Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany) Zoro B. GoneBi (Department of Bioscience, University Felix Houphouet-Boigny, Abidjan, Cote d’Ivoire; Tai Chimpanzee Project, CSRS, Abidjan, Cote d’Ivoire) Tobias Gräßle (Helmholtz Centre for Infection Research, Greifswald, Germany; Robert Koch Institute, Berlin, Germany) Philipp Gunz (Max Planck Institute for Evolutionary Anthropology, Leipzig, Germany) Jennifer E. Jaffe (Robert Koch Institute, Berlin, Germany; Tai Chimpanzee Project, CSRS, Abidjan, Cote d’Ivoire) Anna Jauch (Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany) Fabian H. Leendertz (Helmholtz Institute for One Health, Greifswald, Germany: Robert Koch Institute, Berlin, Germany) Matyas Liptovszky (Twycross Zoo, UK) Patrice Makouloutou Nzassi (Institut de Recherche en Ecologie Tropicale, Libreville, Gabon) Matthew McLennan (Bulindi Chimpanzee and Community Project, School of Social Sciences, Oxford Brooks University, UK) Sophie Moittie (Twycross Zoo, UK) Torsten Møller (Kolmarden Zoo, Sweden) Karin Olofsson-Sannö (National Veterinary Institute, Uppsala, Sweden) Kerrin Pine (Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany) Andrea Pizarro (Tacugama Chimpanzee Sanctuary, Freetown, Sierra Leone) Kamilla Pleh (Robert Koch Institut, Berlin, Germany: Tai Chimpanzee Project, CSRS, Abidjan, Cote d’Ivoire) Jessica Rendel (Twycross Zoo, UK) Liran Samuni (Human Evolutionary Biology, Harvard University, Cambridge, USA; Tai Chimpanzee Project, CSRS, Abidjan, Cote d’Ivoire) Mark Stidworthy (International Zoo Veterinary Group, Keighley, UK) Lara Southern (Institute for Cognitive Sciences, University of Osnabrück, Germany; Ozouga, Loango Chimpanzee Project, Loango National Park, Gabon) Tanguy Tanga (Institut de Recherche en Ecologie Tropicale, Libreville, Gabon; Ozouga, Loango Chimpanzee Project, Loango National Park, Gabon) Steve Unwin (Wildlife Health Australia, Sydney, Australia) Sue Walker (Chester Zoo, UK) Kim Wood (Welsh Mountain Zoo, UK) Klaus Zuberbuehler (Institute of Biology, University of Neuchatel, Switzerland; Budongo Conservation Field Station, Masindi, Uganda)
References
(1) Zucca, F. A.; Segura-Aguilar, J.; Ferrari, E.; Muñoz, P.; Paris, I.; Sulzer, D.; Sarna, T.; Casella, L.; Zecca, L. Interactions of Iron, Dopamine and Neuromelanin Pathways in Brain Aging and Parkinson’s Disease. Prog. Neurobiol.2017, 155, 96–119. https://doi.org/10.1016/j.pneurobio.2015.09.012.
(2) Zecca, L.; Youdim, M. B. H.; Riederer, P.; Connor, J. R.; Crichton, R. R. Iron, Brain Ageing and Neurodegenerative Disorders. Nat. Rev. Neurosci. 2004, 5 (11), 863–873. https://doi.org/10.1038/nrn1537.
(3) Miletić, S.; Bazin, P.-L.; Isherwood, S. J. S.; Keuken, M. C.; Alkemade, A.; Forstmann, B. U. Charting Human Subcortical Maturation across the Adult Lifespan with in Vivo 7 T MRI. NeuroImage 2022, 249, 118872. https://doi.org/10.1016/j.neuroimage.2022.118872.
(4) Langkammer, C.; Krebs, N.; Goessler, W.; Scheurer, E.; Ebner, F.; Yen, K.; Fazekas, F.; Ropele, S. Quantitative MR Imaging of Brain Iron: A Postmortem Validation Study. Radiology 2010. https://doi.org/10.1148/radiol.10100495.
(5) Brammerloh, M.; Morawski, M.; Friedrich, I.; Reinert, T.; Lange, C.; Pelicon, P.; Vavpetič, P.; Jankuhn, S.; Jäger, C.; Alkemade, A.; Balesar, R.; Pine, K.; Gavriilidis, F.; Trampel, R.; Reimer, E.; Arendt, T.; Weiskopf, N.; Kirilina, E. Measuring the Iron Content of Dopaminergic Neurons in Substantia Nigra with MRI Relaxometry. NeuroImage2021, 239, 118255. https://doi.org/10.1016/j.neuroimage.2021.118255.
(6) Gräßle, T.; Crockford, C.; Eichner, C.; Girard‐Buttoz, C.; Jäger, C.; Kirilina, E.; Lipp, I.; Düx, A.; Edwards, L.; Jauch, A.; Kopp, K. S.; Paquette, M.; Pine, K.; EBC Consortium; Haun, D. B. M.; McElreath, R.; Anwander, A.; Gunz, P.; Morawski, M.; Friederici, A. D.; Weiskopf, N.; Leendertz, F. H.; Wittig, R. M. Sourcing High Tissue Quality Brains from Deceased Wild Primates with Known Socio‐ecology. Methods Ecol. Evol. 2023, 2041-210X.14039. https://doi.org/10.1111/2041-210X.14039.
(7) Bazin, P.-L.; Alkemade, A.; Mulder, M. J.; Henry, A. G.; Forstmann, B. U. Multi-Contrast Anatomical Subcortical Structures Parcellation. eLife 2020, 9, e59430. https://doi.org/10.7554/eLife.59430.
(8) Yablonskiy, D. A.; Haacke, E. M. Theory of NMR Signal Behavior in Magnetically Inhomogeneous Tissues: The Static Dephasing Regime. Magn. Reson. Med. 1994, 32 (6), 749–763. https://doi.org/10.1002/mrm.1910320610.
(9) Fedorow, H.; Halliday, G. M.; Rickert, C. H.; Gerlach, M.; Riederer, P.; Double, K. L. Evidence for Specific Phases in the Development of Human Neuromelanin. Neurobiol. Aging 2006, 27 (3), 506–512. https://doi.org/10.1016/j.neurobiolaging.2005.02.015.
Figures
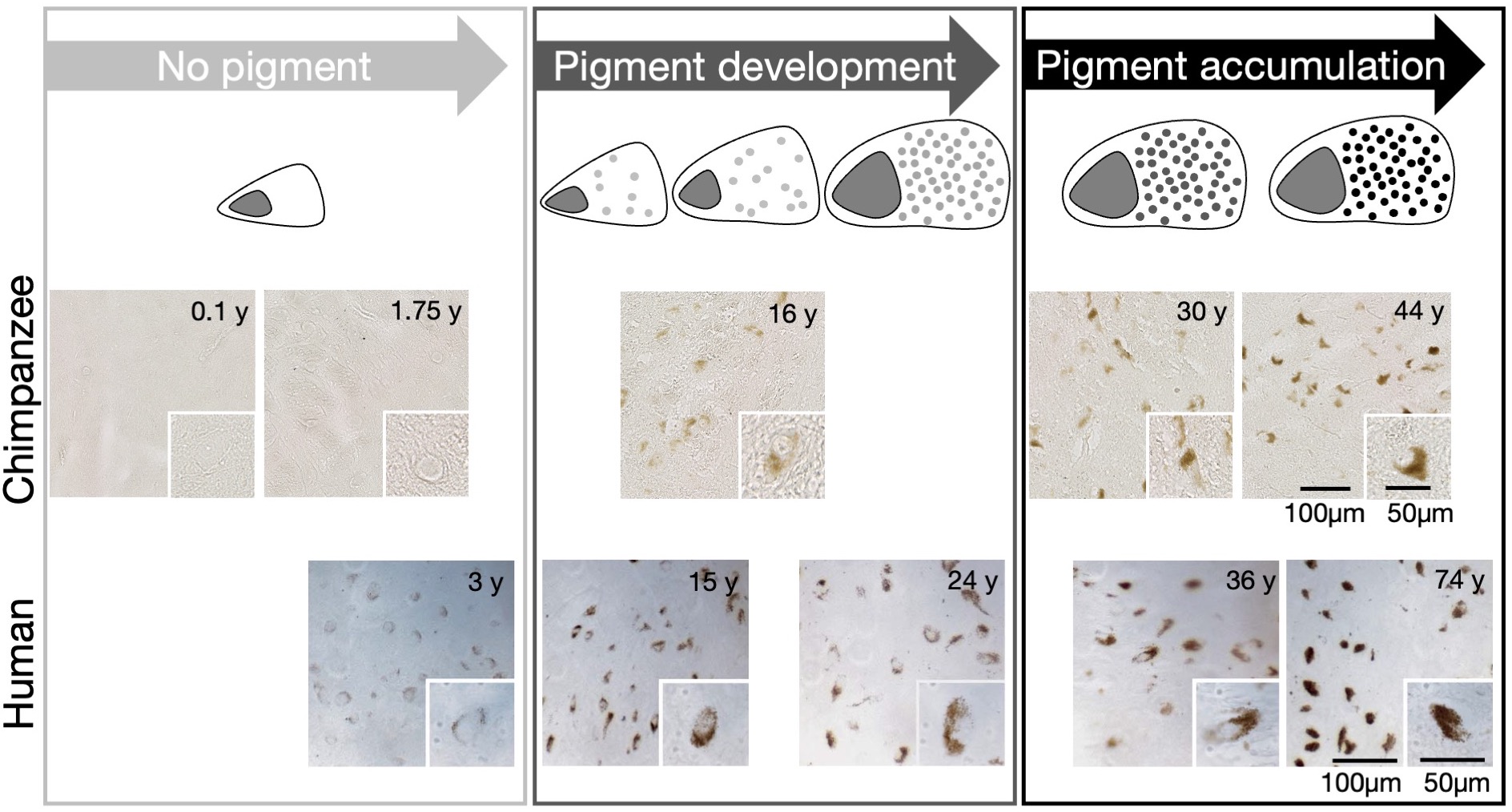
Neuromelanin accumulation in DNs of chimpanzee (upper row, current study) and human midbrains (lower row, adapted from Fedorov9) show the same phases in optical microscopy of unstained slices: No NM until age of 3y, period of pigment development until the age of about 30y, period of pigment accumulation for ages older than 30y9. This qualifies the chimpanzee as an appropriate animal model to study NM accumulation across the human lifespan.