1311
Anisotropy of Longitudinal Relaxation in White Matter: Comparison of T1 and Magnetization Transfer1NMR Methods & Development Group, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany, 2Department of Neurophysics, Max Planck Institute for Human Cognitive and Brain Sciences, Leipzig, Germany, 3Center of Neuropathology and Brain Research, Medical Faculty, University of Leipzig, Paul Flechsig Institute, Leipzig, Germany, 4Felix Bloch Institute for Solid State Physics, Leipzig University, Leipzig, Germany
Synopsis
Keywords: Relaxometry, Relaxometry, T1 Relaxation, Magnetization Transfer, Orientation Dependence
Motivation: Studies on the orientation dependence of T1 have led to contradictory observations, indicating that this effect is not well understood.
Goal(s): Our primary objective was to meticulously explore the orientation dependency of T₁ under precisely controlled and stable conditions.
Approach: Comprehensive T1 (inversion recovery) and magnetization-transfer (MT) experiments were performed in fixed spinal-cord samples, with systematic variation of the fiber-to-field angle θFB .
Results: No relevant T1 variation with θFB was observed in the IR experiments. However, a clear orientation dependence was consistently observed in all MT experiments,
Impact: Precise quantitative MR measurements in spinal cord with varying fiber-to-field angle showed no consistent orientation dependence of T1 but a clear effect for the MT saturation, indicating the MT effects may be responsible for the previously observed T1 anisotropy.
Introduction
White matter tracts are characterized by structural order related to the arrangement of myelin lipid bilayers wrapped around approximately cylindrical axons. Due to anisotropy of residual dipolar couplings for this preferential alignment of lipid molecules, an orientation dependence of MRI contrast may arise, which has been demonstrated in magnetization-transfer (MT) imaging in vivo.1 Studies on orientation dependence of T1 have yielded conflicting observations2-6, including a maximum around fiber-to-field angles of θFB≈50°2,5 or a decrease with increasing θFB3 in human brain in vivo or only minimal variations (1–2%) at room temperature in fresh post-mortem pig brain.4 To better understand such discrepancies, comprehensive T1 and MT experiments were performed on spinal cord samples, where θFB was systematically varied over the full angular range.Methods
Sample preparation: Two pieces of hamster spinal cord (length 27.1 and 24.8 mm, diameter ≤4 mm, Figure 1) were carefully dissected immediately after death and fixated with paraformaldehyde for 3–4 weeks. Subsequently, the cylindrical specimens were washed in phosphate-buffered saline (pH 7.4) and placed in 5mm NMR tubes, embedded in Fomblin.MR experiments: Data were acquired at 3 T (MAGNETOM Sykrafit; Siemens Healthineers) with a custom-made TxRx Helmholtz coil (16 mm radius).7 The coil was mounted inside a wooden box, maintained at approximately constant temperature (36.8–39.0 °C and 35.6–36.6 °C) during 8–9 hours of scanning by a stream of heated air. Continuous fiber-optic temperature monitoring (Optocon; Weidmann) was performed at two positions inside the setup. The sample orientation relative to B0 was remotely adjusted without alteration of the coil properties or removal of the sample. Angles between 0° and 90° were adjusted in randomized order (sample orientation was always at 90° in the first and last scan). Customized pulse sequences with a 1D readout gradient (306 µm nominal resolution) along the sample axis were employed for all acquisitions including the following:
(i) Inversion-recovery (IR) experiments with rectangular inversion (40 µs) and readout pulses (20 µs) and 23 logarithmically spaced inversion times (TI between 770 μs and 10 s, TR=13 s, TE=7.5 ms).
(ii) Measurements of MT saturation (MTsat = 1 - MTR = S(MT)/S0) with a train of Gaussian RF pulses (n=250, duration 2 ms, separation 250 µs, offset frequency 15 kHz, root-mean-square amplitude 2π·500 rad/s), which were either applied single-sided or dual-sided.
(iii) Diffusion-tensor imaging (DTI) with 60 gradient directions (TE=80 ms, b=1500 s/mm2) to reconstruct the local average fiber direction and fractional anisotropy (FA).
Results and Discussion
The DTI results indicated moderate to high anisotropy in both specimens (Figure 2). Two different ROIs (approx. 10 voxels) were chosen in each sample for further analyses, one with FA>0.65 (indicated in blue) and one of somewhat lower FA (0.47–0.51; indicated in orange).Orientation dependence of MT-induced signal change: In all experiments, including single-sided (Figure 3) and dual-sided application of off-resonance saturation pulses (Figure 4), a clear orientation dependence was consistently observed for MT saturation. The pattern of variation (order of 5–10% with a maximum between θFB≈30–40°) was similar to that reported for the apparent T2 of the semisolid pool in previous quantitative MT experiments in vivo.1 As expected, the variations were generally enhanced in ROIs of higher FA.
Orientation dependence of T1: Despite a high degree of orientational order, no consistent pattern of T1 variation with θFB was observed in the IR experiments (Figure 4). This is in contrast to previous human studies reporting detectable changes for FA>0.3.8 T1 variations were <2% (average values around 690 ms), which is of the order of the experimental accuracy. Additional measurements yielded a temperature-related T1 change by 4 ms/K. Hence, contributions to the subtle T1 variations from temperature drifts (order of ±2 K) cannot be completely excluded.
Conclusion
The characteristic alterations of MTR with θFB are probably due to distinct changes of the underlying dipolar lineshape of the macromolecular pool.1 Remarkably, our results obtained under carefully controlled conditions and with high SNR do not support the assumption of a relevant (direct) orientation dependence of T1 of the water pool. Notably, the hard pulses used in our 1D IR experiments achieve a well-defined preparation of the spin system (water and macromolecules), whereas previous observations of orientation effects on T1 were obtained with MP2RAGE2 or variable-flip-angle imaging techniques,3 comparing results in separate white-matter ROIs with different main fiber orientation without sample rotations. These methods involve the application of a large number of soft readout pulses, which may produce spatially variable MT weighting and, thereby, impact (indirectly) the recovery of the water magnetization.Acknowledgements
No acknowledgements found.References
- Pampel A, Müller DK, Anwander A, Marschner H, Möller HE. Orientation dependence of magnetization transfer parameters in human white matter. NeuroImage 2015; 114: 136–146.
- Kauppinen RA, Thotland J, Pisharady PK, Lenglet C, Garwood M. White matter microstructure and longitudinal relaxation time anisotropy in human brain at 3 and 7 T. NMR Biomed. 2023; 36: e4815.
- Schyboll F, Jaekel U, Weber B, Neeb H. The impact of fibre orientation on -relaxation and apparent tissue water content in white matter. Magn. Reson. Mater. Phy. 2018; 31(4): 501-510.
- Wallstein N, Pampel A, Jäger C, Müller R, Möller HE. Investigations of T1 anisotropy in ex-vivo white matter using a tiltable coil. Proceedings of the 31st Annual Meeting of ISMRM, Toronto, ON, Canada, 2023. p. 1358.
- Kauppinen RA, Thothard J, Leskinen HPP, Pisharady PK, Manninen E, Kettunen M, Lenglet C, Grohn OHJ, Garwood M, Nissi MJ. Axon fiber orientation as the source of T1 relaxation anisotropy in white matter: A study on corpus callosum in vivo and ex vivo. Magn. Reson. Med. 2023; 90: 708–721.
- Henkelman RM, Stanisz GJ, Kim JK, Bronskill MJ. Anisotropy of NMR properties of tissues. Magn. Reson. Med. 1994; 32: 592–601.
- Wallstein N, Pampel A, Müller R, Möller HE. Radiation damping at clinical field strength: Characterization and compensation in quantitative measurements. Magn. Reson. Med. (in press) DOI: 10.1002/MRM.29934.
- Knight MJ, Damion RA, Kauppinen RA. Observation of angular dependence of T1 in the human white matter at 3T. Biomed. Spectrosc. Imaging 2018; 7: 125–133.
Figures
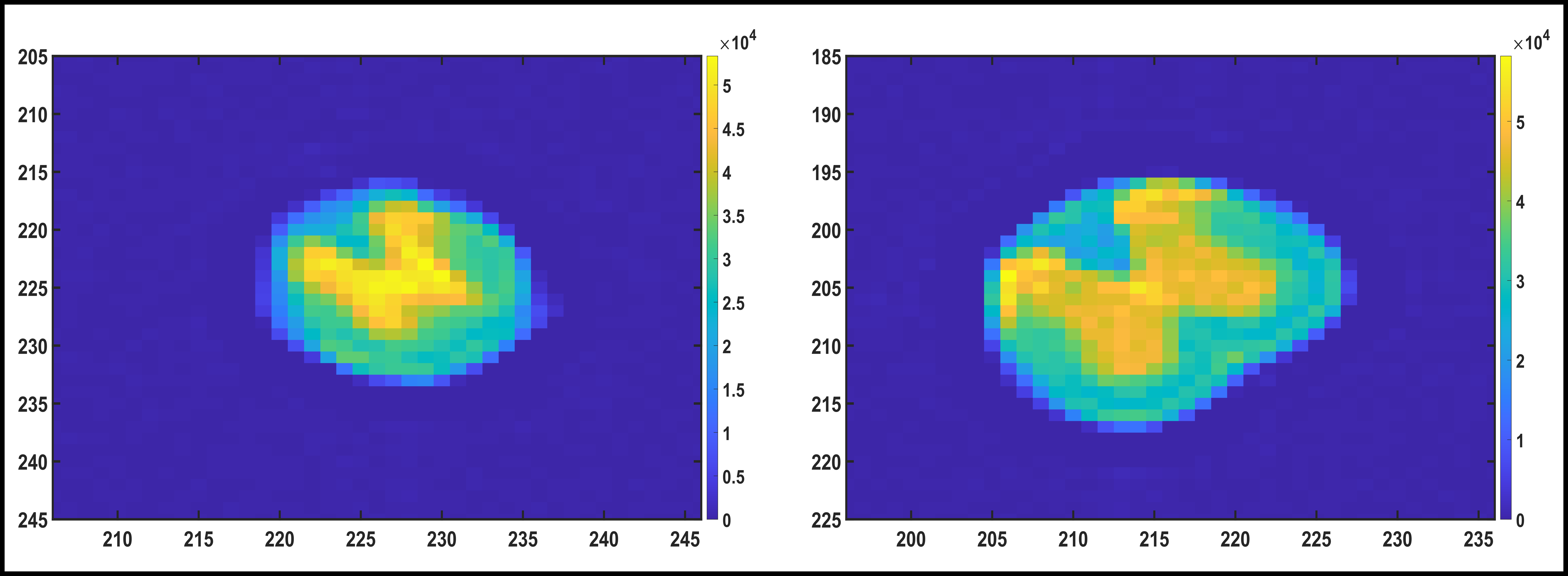
FIGURE 1: Cross-section through one of the spinal cord samples (thinner part on the left vs. thicker part on the right, in-plane resolution 125 μm, slice thickness 3 mm) demonstrating the characteristic arrangement of gray matter (central part) and white matter (periphery). While related partial-volume effects in 1D experiments constitute a potential limitation, the average FA values (see Figure 2) indicated a sufficiently high degree of orientational order.
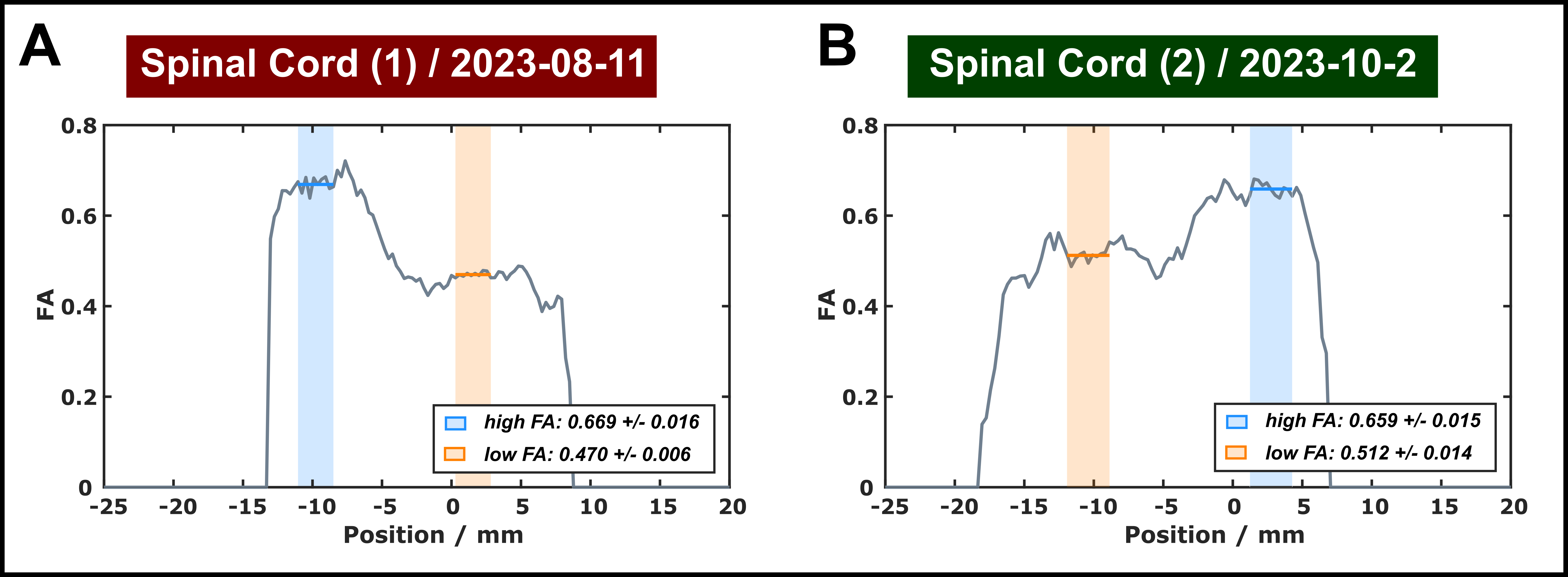
FIGURE 2: Results of 1D DTI in both samples. Blue and orange shading indicate ROIs selected for the further analysis with average FA values of 0.66–0.67 and 0.47–0.51, respectively.
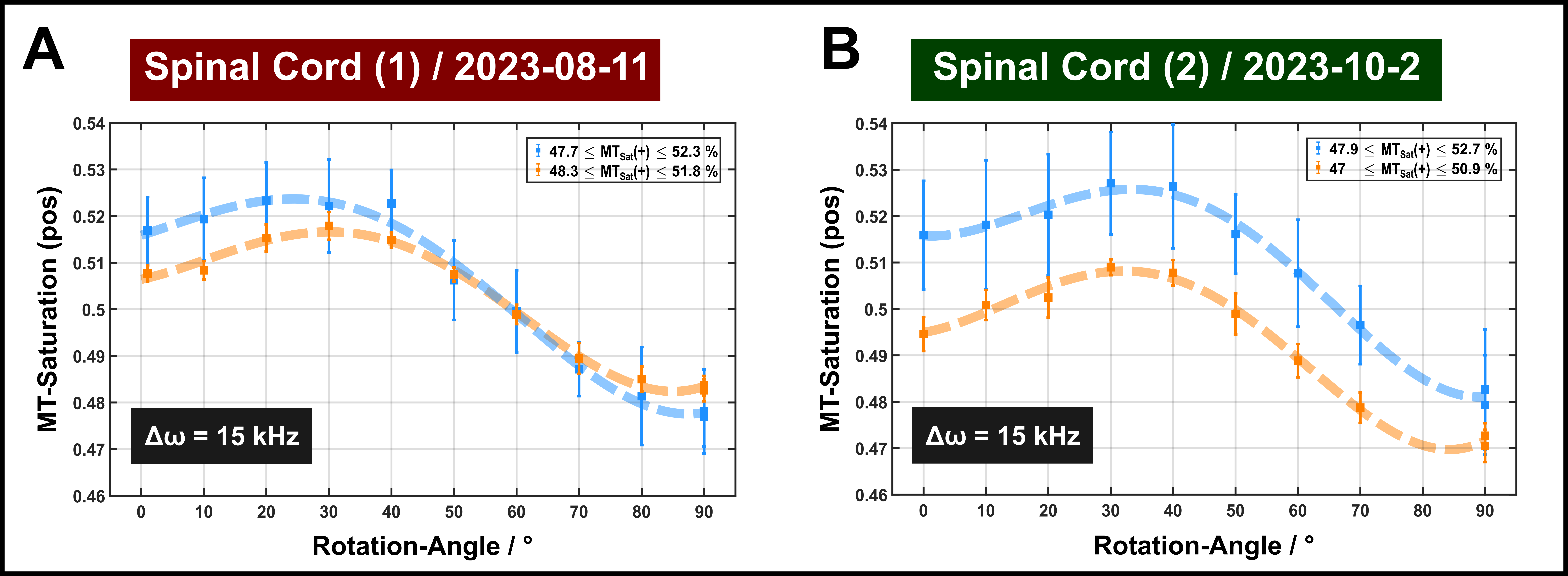
FIGURE 3: MT saturation measurements in the two ROIs (blue and orange symbols consistent to Figure 2 with dashed lines guiding the eye) obtained with a single-sided off-resonant saturation with an offset-frequencies of 15 kHz. Error bars indicate standard deviations across the ROI.
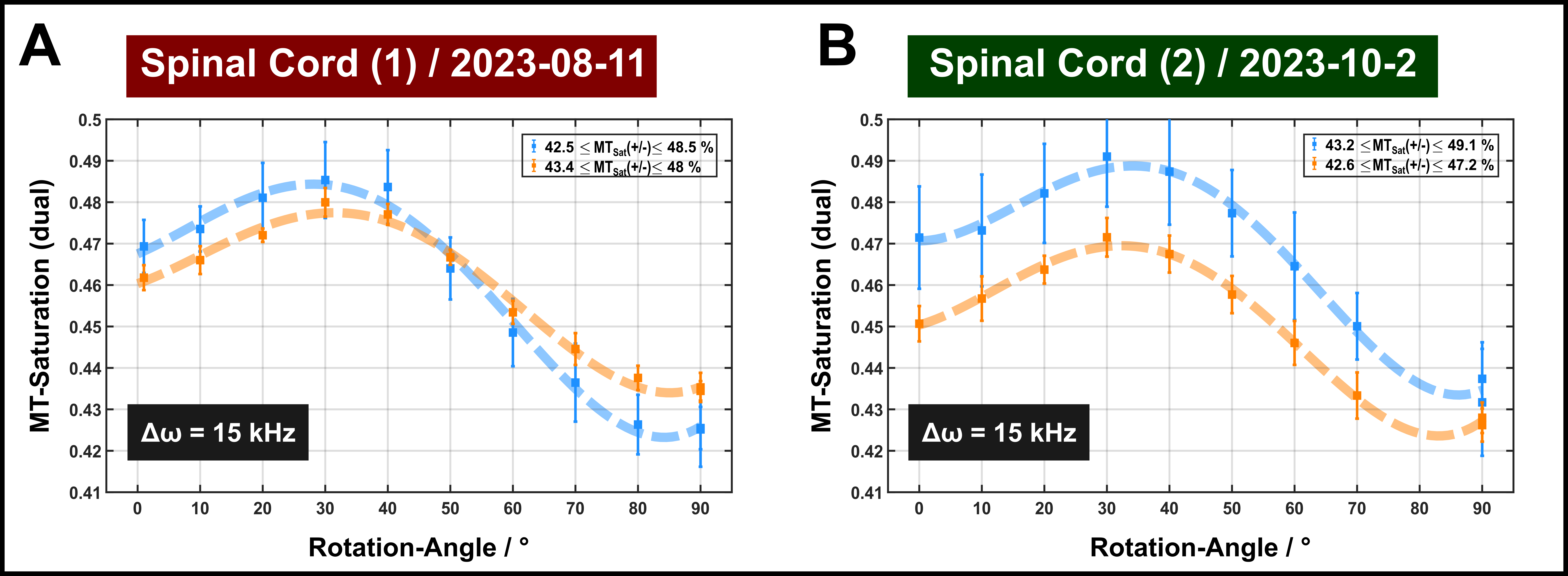
FIGURE 4: MT saturation measurements in the two ROIs (blue and orange symbols consistent to Figure 2 with dashed lines guiding the eye) obtained with a dual-sided off-resonant saturation with an offset-frequencies of 15 kHz. Error bars indicate standard deviations across the ROI.
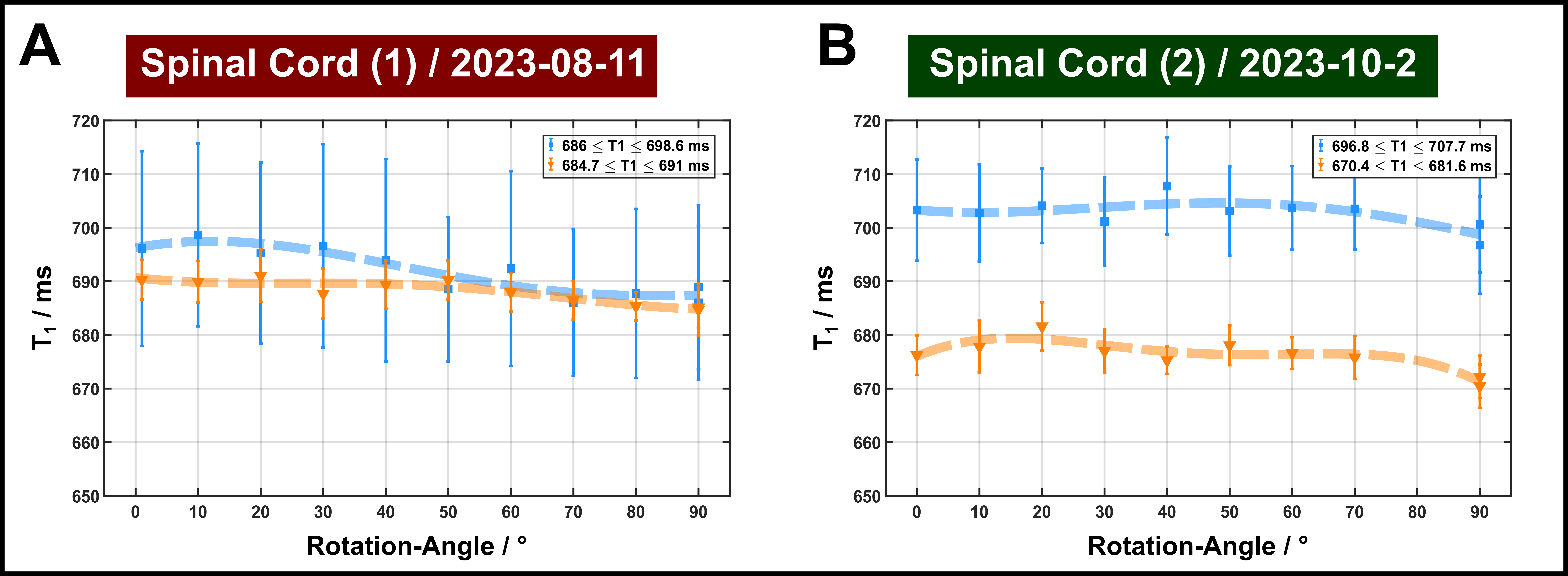
FIGURE 5: T1 relaxation times (blue and orange symbols consistent to Figure 2 with dashed lines guiding the eye) obtained with inversion recovery assuming mono-exponential behavior for inversion times >100 ms, as typically employed in in-vivo experiments. Error bars indicate standard deviations across the ROI.