1154
Neurological Underpinning of Cortical Folding in the Human Fetal Brain Using In-utero MRI-Informed Computational Modeling1Key Laboratory for Biomedical Engineering of Ministry of Education, Department of Biomedical Engineering, College of Biomedical Engineering & Instrument Science, Zhejiang University., Hangzhou, China
Synopsis
Keywords: Fetal, Diffusion/other diffusion imaging techniques, Brain, Simulation
Motivation: Computational models may help to decipher the neuroscientific mechanisms of cortical folding. Past simulations relied on histology or ex-vivo MRI data, which may not fully capture the complexity of in-utero brain development.
Goal(s): To build a simulation model leveraging in-utero MRI data to elucidate how cortical microstructures impact fetal brain gyrification.
Approach: Based on diffusion MRI-measured microstructures, we simulated cortical folding in the right temporal lobe using a computational model.
Results: Differences in fiber density between sulci and gyri are critical for folding initialization and development, linked to regional differences in dendritic arborization. Simulation results agreed with experimentally measured fetal brain morphology.
Impact: Our work introduced a novel computational model that utilized in-utero MRI data to simulate the cortical folding process of the human fetal brain, suggesting local differences in dendritic arborization may be one of the driving forces of cortical folding.
Introduction
Cortical folding is a complex process involving various cellular and mechanical activities, but its exact drivers remain largely unknown 1-3. One leading theory, “differential growth”, suggests the higher growth rate in the cortical plate (CP) causes brain surface buckling 4. However, previous simulations of this theory failed to represent the realistic patterns of the human fetal brain, and did not consider the physical information from in-utero measurements 1,5-7.We previously generated a spatiotemporal atlas of normal developing fetal brains using in-utero T2 weighted (T2w) and diffusion MRI (dMRI) between 23-38 gestational weeks (W) 8,9. In this study, we combined these data with mechanical modeling to simulate fetal brain cortical folding in the temporal region. Our simulations closely matched actual cortical morphology. Notably, the fiber fraction from diffusion basis spectrum imaging (DBSI) played a key role in our simulation, indicating cortical dendritic arborization may be critical for folding generation and development.
Material and Methods
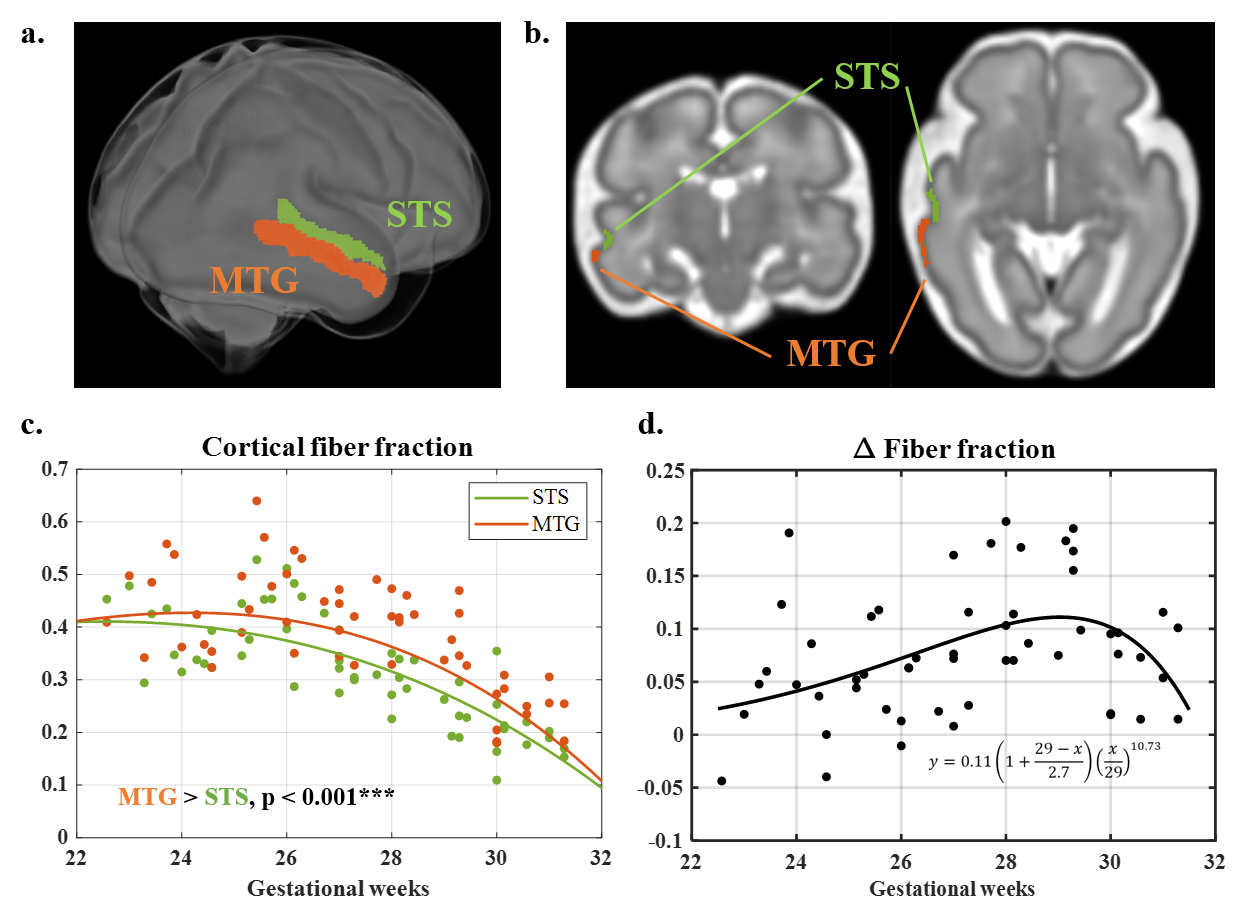
We utilized spatiotemporal in-utero T2w and dMRI atlases of fetal brains between 23W to 31W 8,9, containing T2w images and segmentation labels, and diffusion tensor imaging (DTI) based fractional anisotropy (FA) and apparent diffusivity coefficient (ADC). Additionally, DBSI model was used to calculate the fractions of the fiber, restricted, hindered, and restricted components 10.Our study focused on one of the earliest developing folds. We manually delineated the superior temporal sulcus (STS) and middle temporal gyrus (MTG) for each week (Figure 3b), which were transformed into subject spaces. Fiber fraction was extracted from subjects and compared between STS and MTG using paired t-test. The difference in fiber fraction between the sulcus and the gyrus was fitted to a growth-and-decline model, resulting in a function df 11.
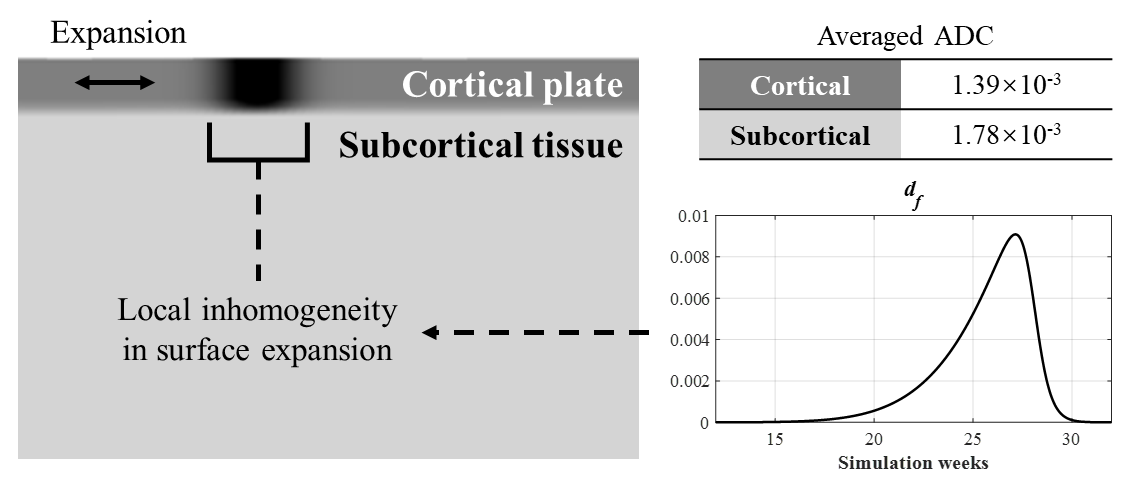
For our simulation in COMSOL Multiphysics version 5.3a, we designed a 2D model replicating the initial shape near STS and MTG. The CP and subcortical tissue were defined as separated compartments (Figure 1), with shear modulus of 300Pa and 234Pa respectively, according to previous definitions and the ADC ratio between them 1,6. In continuum mechanics, the total deformation can be formulated as
F = Fe·G (1)
where Fe is the elastic deformation tensor and G is the growth tensor, with both tangential and radial growth components. We assumed no radial growth and the tangential growth is an exponential function according to our previous study 8:
Gy(t) = 1, Gx(t) = egct (2)
Where the tangential growth rate gc was 0.01 in the temporal lobe in 2D. Subcortical tissue has no growth but only an elastic response. We assumed a local increase in gc in MTG induced by extensive dendritic arborization, quantified by df and a response ratio α:
g*c = gc + αdf (3)
The time-dependent model was solved numerically using finite-element methods. At each time point, the output morphology was displayed and the mean absolute curvature was calculated.
Results
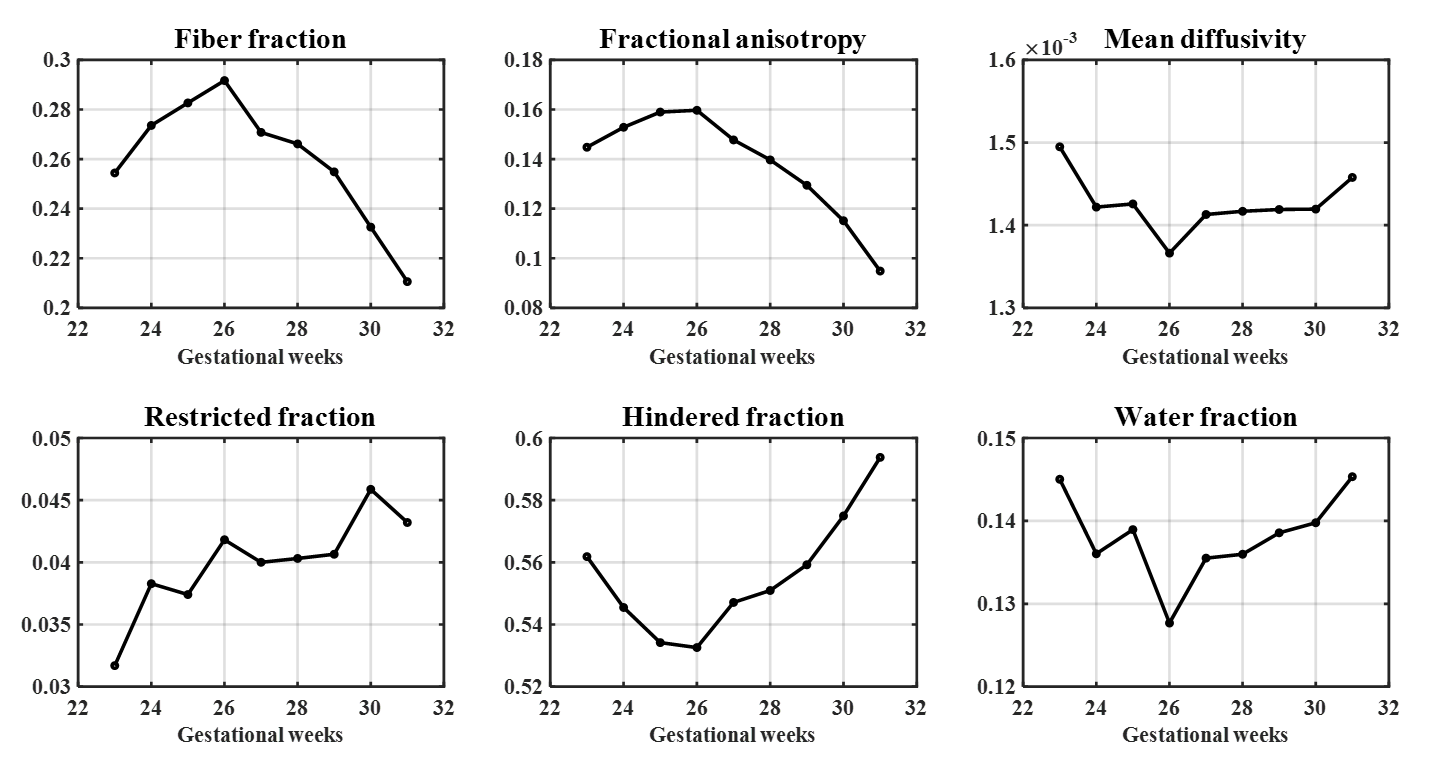
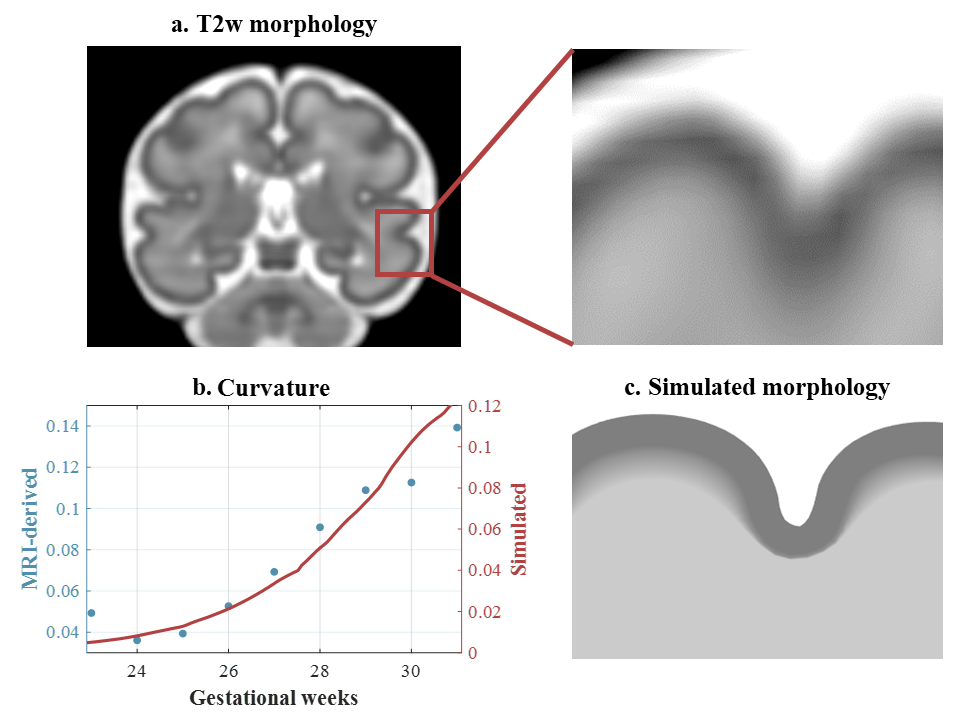
In the whole-brain CP, the restricted fraction increased from 23W to 31W. Before 26W, FA and fiber fraction increased while diffusivity, hindered and water fraction decreased, suggesting radial neuronal migration and axonal ingrowth. After 26W, FA and fiber fraction declined while hindered and water fraction increased. These patterns might result from the loss of radial glial cells, increased extra-cellular matrix, and dendritic arborization.Notably, fiber fraction was significantly higher in MTG than in STS (t = 7.79, p < 0.001), and their difference df displayed a non-monotonic trend (Figure 3). Fiber fraction is known to correlate with dendrite density 12, and the change in dendritic components had a close correlation to cortical curvature 3. Accordingly, we used df to quantify local differences in cortical growth during simulation (Eq. 3). The simulated 2D model resembled the folding pattern of MTG and STS at 31W (Figure 4a,c). The simulated curvature displayed similar trends to MRI-derived curvature (Figure 4b).
Discussion
In this study, we observed complex changes in cortical DTI and DBSI quantifications from 23W to 31W, indicating diverse roles of the microstructural developments. Specifically, the change in DBSI-based fiber fraction may reflect the role of dendritic arborization in driving cortical folding, as demonstrated by our computational model. The MRI-informed mechanical model was able to resemble the real-world development of cortical folding.Future research should consider additional biological factors for a more realistic model, such as changes in tissue stiffness, cortical thickness during development, and the brain's initial morphology. Furthermore, simulations on other gyri and sulci should be performed to validate our assumptions.
Conclusion
Our work incorporated in-utero MRI data into mechanical modeling to perform computational simulation, shedding light on the mechanism of cortical folding.Acknowledgements
This work was supported by the National Natural Science Foundation of China (81971606, 82122032), and the Science and Technology Department of Zhejiang Province (202006140, 2022C03057).References
1. Xu G, Knutsen A K, Dikranian K, et al. Axons Pull on the Brain, But Tension Does Not Drive Cortical Folding. Journal of Biomechanical Engineering-Transactions of the Asme. 2010;132(7).
2. Reillo I, de Juan Romero C, Garcia-Cabezas M A, et al. A Role for Intermediate Radial Glia in the Tangential Expansion of the Mammalian Cerebral Cortex. Cerebral Cortex. 2011;21(7):1674-1694.
3. Wang X, Studholme C, Grigsby P L, et al. Folding, But Not Surface Area Expansion, Is Associated with Cellular Morphological Maturation in the Fetal Cerebral Cortex. Journal of Neuroscience. 2017;37(8):1971-1983.
4. Richman D P, Stewart R M, Hutchinson J W, et al. MECHANICAL MODEL OF BRAIN CONVOLUTIONAL DEVELOPMENT. Science. 1975;189(4196):18-21.
5. Tallinen T, Chung J Y, Rousseau F, et al. On the growth and form of cortical convolutions. Nature Physics. 2016;12(6):588-593.
6. Garcia K E, Wang X and Kroenke C D. A model of tension-induced fiber growth predicts white matter organization during brain folding. Nature Communications. 2021;12(1).
7. Wang S, Saito K, Kawasaki H, et al. Orchestrated neuronal migration and cortical folding: A computational and experimental study. Plos Computational Biology. 2022;18(6).
8. Xu X, Sun C, Sun J, et al. Spatiotemporal atlas of the fetal brain depicts cortical developmental gradient in Chinese population. bioRxiv. 2022.
9. Chen R, Sun C, Liu T, et al. Deciphering the developmental order and microstructural patterns of early white matter pathways in a diffusion MRI based fetal brain atlas. NeuroImage. 2022;119700-119700.
10. Spees W M, Lin T-H, Sun P, et al. MRI-based assessment of function and dysfunction in myelinated axons. Proceedings of the National Academy of Sciences of the United States of America. 2018;115(43):E10225-E10234.
11. Yin X Y, Goudriaan J, Lantinga E A, et al. A flexible sigmoid function of determinate growth. Annals of Botany. 2003;91(6):316-370.
12. Zhan J, Lin T-H, Libbey J E, et al. Diffusion Basis Spectrum and Diffusion Tensor Imaging Detect Hippocampal Inflammation and Dendritic Injury in a Virus-Induced Mouse Model of Epilepsy. Frontiers in Neuroscience. 2018;12.
Figures