1140
Diffusion MRI-based Estimation of Cortical Architecture via Machine Learning (DECAM) enhanced by cortical label vectors1Department of Radiology, Children's Hospital of Philadelphia, Philadelphia, PA, United States, 2Department of Bioengineering, University of Pennsylvania, Philadelphia, PA, United States, 3Department of Radiology, University of Pennsylvania, Philadelphia, PA, United States, 4School of Information Science and Engineering, Dalian University of Technology, Dalian, China
Synopsis
Keywords: Microstructure, Diffusion/other diffusion imaging techniques, Diffusion analysis and visualization, biomarkers, cortical architecture, non-invasive virtual histology
Motivation: Advanced diffusion MRI (dMRI) has enabled noninvasive assessment of cortical measures conventionally only available from neuropathology. Analytical dMRI models are limited by restrictive model assumptions.
Goal(s): In this study, we develop Diffusion-MRI based Estimation of Cortical Architecture using Machine-learning (DECAM), a translational framework of “noninvasive neuropathology” that can quantify cortical architecture based on dMRI.
Approach: DECAM incorporates cortical label vectors to address the challenge of achieving perfect MRI-histology registration in primate brains due to their complex morphology.
Results: By providing high-fidelity, reproducible whole-brain soma density maps validated with histology, DECAM paves the way for data-driven noninvasive histology for potential applications such as Alzheimer’s.
Impact: DECAM is the first translational framework and robust pipeline that addresses the challenge of estimating high-fidelity whole-brain soma density in primate brains with complex morphology. DECAM paves the way for data-driven noninvasive histology for potential applications such as Alzheimer’s.
Purpose
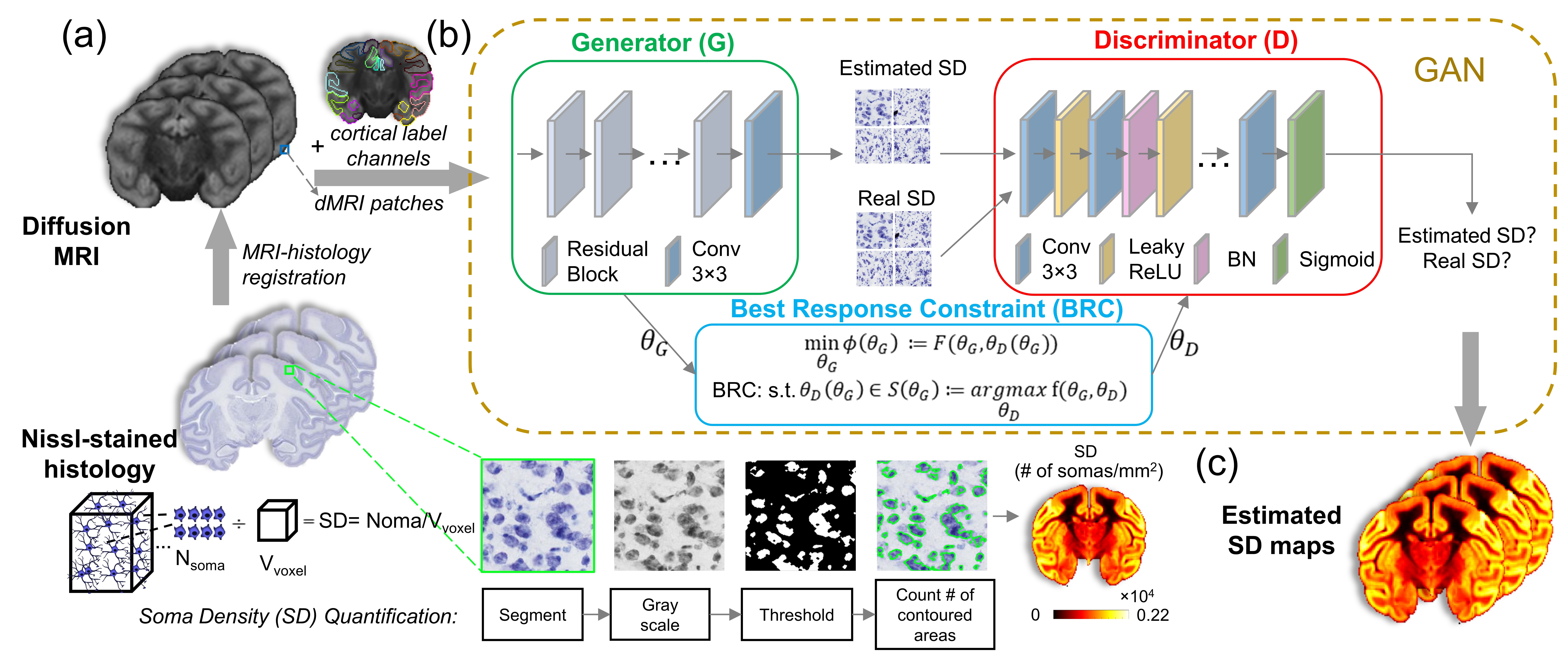
Advanced diffusion MRI (dMRI) has enabled noninvasive assessment of conventional cortical histological measures1-9. However, analytical models are limited by their restrictive model assumptions and lack of validation from quantitative histology. DECAM is a translational framework and robust pipeline that addresses the challenge of estimating high-fidelity whole-brain soma density (SD) in primate brains with complex morphology10. To account for misregistration due to the complex morphology of primate brains, we incorporated cortical label vectors11 into our deep learning (DL) algorithm. By providing high-fidelity, reproducible whole-brain SD maps validated with histology, DECAM paves the way for data-driven, noninvasive virtual histology for potential applications such as Alzheimer’s disease.Methods
DMRI for macaque (Fig. 1a)DMRI acquisitions with two b-values (b=1500, 4500s/mm2) and 30 gradient directions were performed on a normal postmortem Rhesus macaque brain with in-plane resolution 0.6×0.6mm2, slice thickness=2mm, number of averages=24.
Quantification of soma density from histological images
For measuring SD (Fig. 1a bottom), the Nissl-stained histology images of resolution 0.46µm/pixel12 were blocked into segments with size of 0.6×0.6 mm2. Segments are gray-scaled and threshold. SD is defined as number of contoured areas/ segment area in mm2. The calculated SD map agrees well with SD from histology13.
Histology-MRI registration (Fig 1a)
12 Nissl histology slices were gray-scaled and affine registered to average diffusion weighted image (aDWI). SD map was affine registered to aDWI using the same transformation. The registered dMRI volumes and SD maps were cut to 3 by 3 patches and served as training data.
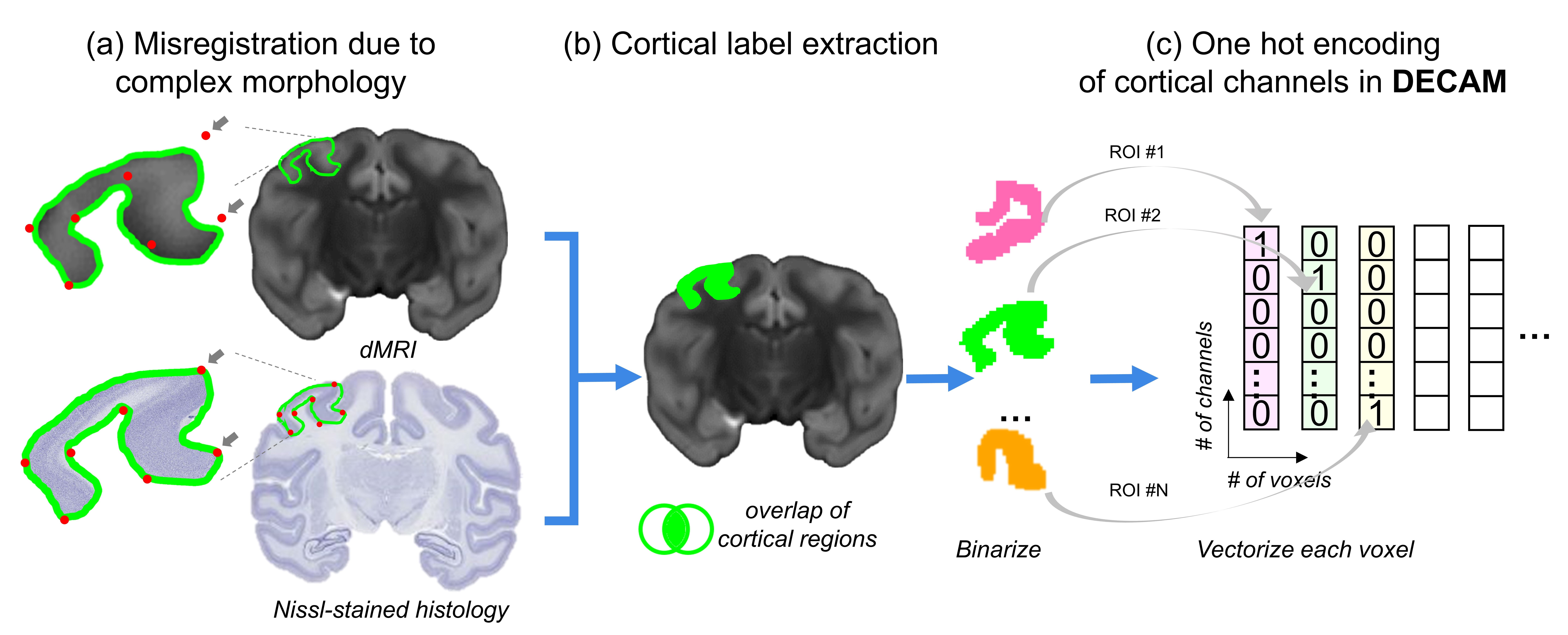
DECAM cortical labels to correct for residual misregistration
After the linear and nonlinear transformations (Fig. 1a), visible residual misregistration remains (Fig. 2a). To resolve the challenge of residual misregistration due to the complex brain morphology of primate brains, we introduced DECAM cortical labels vectors transferred from a macaque atlas14. We found the overlap of cortical labels in dMRI image and in histology image for each cortical region to represent the location of well-registered voxels. We designed N=66 cortical regions of interests (ROIs) to be image volumes with 66 channels, where each channel is a binary mask specifying the spatial domain of a particular structure (Fig. 2c). The cortical labels in 66 channels were fed into the DL network as inputs along with the dMRI volumes in patches (Fig. 2c). The cortical labels ensure that only voxels correctly registered to the same cortical region from histology to dMRI space are used for training the DL algorithm.
Training Best Response Constraint (BRC) Generative Adversarial Network (GAN)
We implemented a Wasserstein GAN with gradient penalty (WGAN-GP) with the BRC15 and trained the model with an Adam optimizer with beta=0.9999 and learning rate=0.0002 for 40 epochs. A mean-squared-error cost function between predicted images and ground-truth was added to the generator cost function to aid prediction.
Estimation of soma density
Whole brain SD was estimated on two separate macaque subjects not used for training the network, and SD was also predicted for a held-out slice on the training macaque brain.
Results
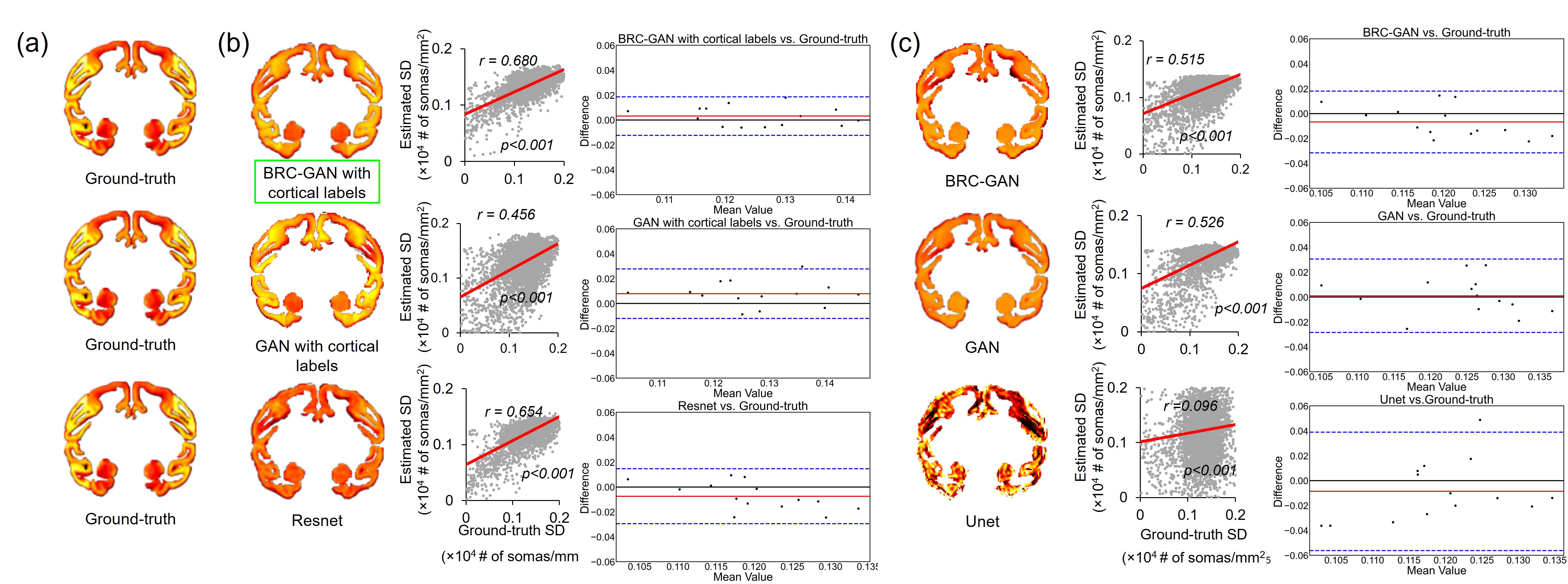
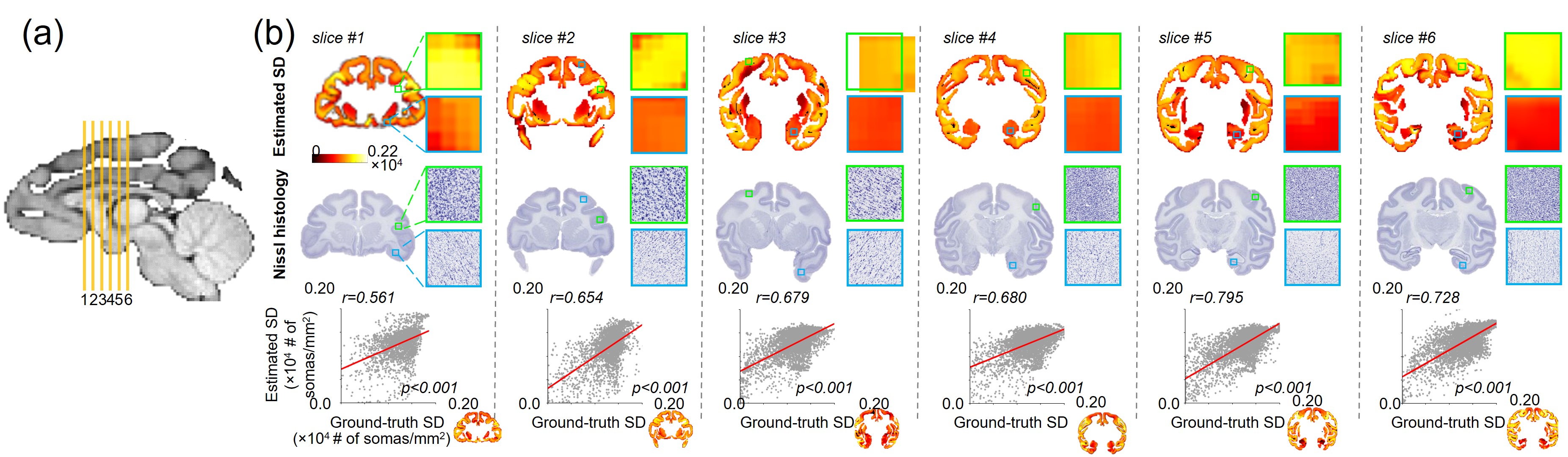
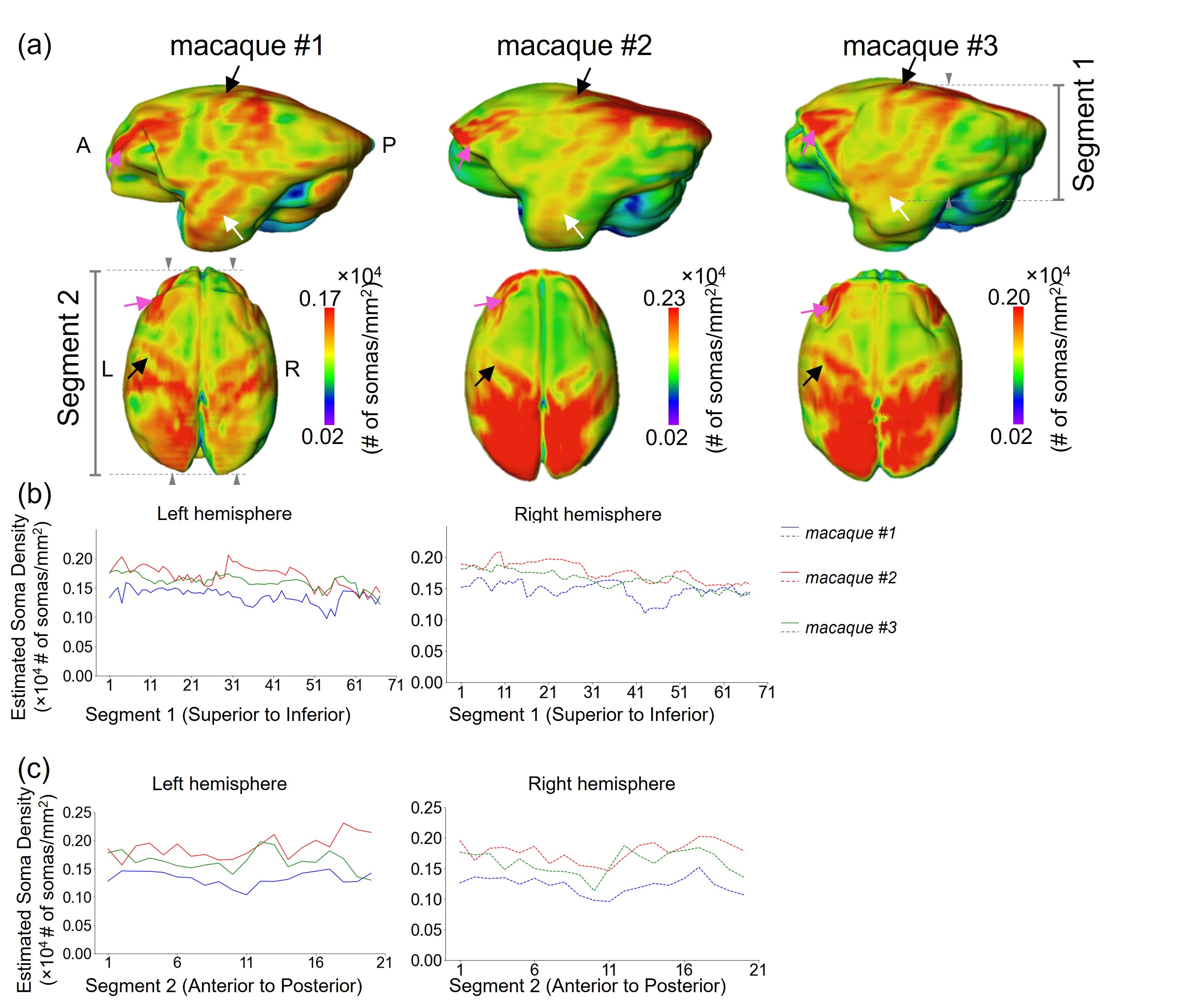
The proposed BRC-GAN with cortical labels achieved the highest correlation (r=0.68) between estimated and ground-truth SD on a held-out test slice across all methods (Fig.3 b,c middle column). On the Blant-Altman plot (Fig. 3 b,c right column) for comparing the difference between estimation from considered DL network and ground-truth SD, the proposed BRC-GAN with cortical labels achieved the lowest bias (red line) and narrowest limits of agreement (blue dotted lines). The variation of estimated SD over the cortex can be clearly observed from the top row of Fig. 4b, while consistency with the corresponding histology slices can be well appreciated from the middle row of Fig. 4b. On all test slices, estimated SD has high correlation (all r values >0.6, p values < 0.001) with the ground-truth SD. Whole brain SD maps were estimated on two additional test macaque brains (macaque #2-3) (Fig. 5a). Overall, the estimated SD maps demonstrate high heterogeneity across the entire cortical surface, with the precentral gyrus (black arrows), the superior frontal gyrus (pink arrows), and the superior temporal gyrus (white arrows) exhibiting consistently higher SD values across subjects (Fig. 5a). The heterogeneous spatial profile is consistent across three subjects (Fig. 5 b,c).Discussion and conclusion
We qualitatively and quantitatively demonstrate the effectiveness of cortical label vectors in DECAM. We also showcase high correspondence between the DECAM-estimated SD maps and ground-truth in macaques and high reproducibility across subjects for whole-brain cortical SD maps. By providing high-fidelity, reproducible estimated SD maps validated with histology, DECAM paves the way for paradigm-shifting data-driven noninvasive virtual histology for potential applications such as Alzheimer’s diseases.Acknowledgements
This study is funded by NIH R01MH092535, R01MH125333, R01EB031284, R01MH129981, R21EB009545, R21MH123930, UM1MH130991 and P50HD105354.References
1. Stanisz GJ, Wright GA, Henkelman RM, Szafer A. An analytical model of restricted diffusion in bovine optic nerve Magn Reson Med. . 1997;37:103-111.
2. Jensen JH, Helpern JA, Ramani A, Lu H, Kaczynski K. Diffusional kurtosis imaging: the quantification of non‐gaussian water diffusion by means of magnetic resonance imaging. Magn Reson Med. 2005;53:1432-1440.
3. Zhou XJ, Gao Q, Abdullah O, Magin RL. Studies of anomalous diffusion in the human brain using fractional order calculus. Magn Reson Med. 2010;63:562-569.
4. Jespersen SN, Kroenke CD, Østergaard L, Ackerman JJ, Yablonskiy DA. Modeling dendrite density from magnetic resonance diffusion measurements. NeuroImage. 2007;34:1473-1486.
5. Zhang H, Schneider T, Wheeler-Kingshott CA, Alexander DC. NODDI: practical in vivo neurite orientation dispersion and density imaging of the human brain. NeuroImage. 2012;61:1000-1016.
6. Le Bihan D, Johansen-Berg H. Diffusion MRI at 25: exploring brain tissue structure and function. NeuroImage. 2012;61:324-341.
7. Alexander DC, Dyrby TB, Nilsson M, Zhang H. Imaging brain microstructure with diffusion MRI: practicality and applications. NMR Biomed. 2019;32:e3841.
8. Novikov DS, Fieremans E, Jespersen SN, Kiselev VG. Quantifying brain microstructure with diffusion MRI: Theory and parameter estimation. NMR Biomed. 2019;32:e3998.
9. Zhu T, Peng Q, Ouyang A, Huang H. Neuroanatomical underpinning of diffusion kurtosis measurements in the cerebral cortex of healthy macaque brains. Magn Reason Med. 2021; 85:1895-1908.
10. Zhu T, Ouyang M, Liu X, Liu R, Huang H Best Response Constraint Generative Adversarial Network for Diffusion MRI-based Estimation of Cortical micro-Architecture. ISMRM2023 abstract #4481
11. Balakrishnan G, Zhao A, Sabuncu MR, Guttag J, Dalca AV: Voxelmorph: a learning framework for deformable medical image registration. IEEE Trans. Med. Imaging. 2019 38(8), 1788-1800
12. Mikula S, Trotts I, Stone JM, Jones EG. Internet-enabled high-resolution brain mapping and virtual microscopy. 2007; NeuroImage. ;35:9-15
13. Carlo CN, Charles FS. Structural uniformity of neocortex, revisited.2013; PNAS. 110.4: 1488-1493.
14. Feng L, Jeon T, Yu Q, Ouyang M, Peng Q, Mishra V, Huang H. Population-averaged macaque brain atlas with high-resolution ex vivo DTI integrated into in vivo space. Brain Structure and Function (2017) 222, 4131-4147.
15. Liu R, Gao J, Liu X, Fan X Revisiting GANs by Best-Response Constraint: Perspective, Methodology, and Application. 2022; arXiv preprint arXiv:2205.10146.
Figures