0971
Dynamic Causal Modeling Reveals Distinct Network-Specific Effective Connectivity within Olfactory Pathway between Healthy and Aged Rats1Laboratory of Biomedical Imaging and Signal Processing, The University of Hong Kong, Hong Kong SAR, China, 2Department of Electrical and Electronic Engineering, The University of Hong Kong, Hong Kong SAR, China, 3Department of Diagnostic Radiology, The University of Hong Kong, Hong Kong SAR, China, 4School of Chinese Medicine, The University of Hong Kong, Hong Kong SAR, China, 5School of Biomedical Sciences, The University of Hong Kong, Hong Kong SAR, China
Synopsis
Keywords: Aging, Brain Connectivity, fMRI Analysis, functional connectivity, neuroscience, fMRI (task-based)
Motivation: Presently, olfactory dysfunctions such as with aging, neurodegenerative diseases and COVID-19 remain poorly understood at the systems level despite extensive knowledge of the microcircuit changes at the olfactory bulb (OB).
Goal(s): We aim to reveal the systematic abnormalities of downstream olfactory information processing from the OB in prematurely aged rats.
Approach: We examined the effective connectivity of olfactory networks in both healthy and aged rat models with optogenetic fMRI and dynamic causal modeling.
Results: We demonstrate that network-specific dynamics in the olfactory system between aged and healthy rats could be attributed to altered effective connectivity driven by primary olfactory regions downstream from OB.
Impact: The ability to stimulate olfactory bulb excitatory neurons and model the downstream neural activity dynamics at the system level in healthy and aged animals has revealed key regions that are involved in olfactory dysfunctions, which can guide future therapeutic interventions.
Purpose
Decreased olfactory function with aging parallels generalized age-related deficits in sensory functions and cognition that occur in the absence of obvious disease states1. However, studies in rodents have shown that the decreased ability to perceive odors was not associated with a general decline in the number of receptors/cells in the olfactory epithelium or a decline in neuronal populations (i.e., mitral and tufted cells, and interneurons) in the olfactory bulb (OB)2. Instead, the synaptic density in the glomeruli of the OB appears diminished2, suggesting impaired downstream olfactory information processing from the OB to targets in the subcortical and cortical regions.Here, we propose to examine the spatiotemporal response property and the effective connectivity of key regions within the olfactory network in an aging rat model with optogenetic fMRI and dynamic causal modeling (DCM). We aim to provide insights into the systematic abnormalities of downstream olfactory information processing from the OB, specifically the network-specific effective connectivity changes in both local and long-range olfactory circuits.
Methods
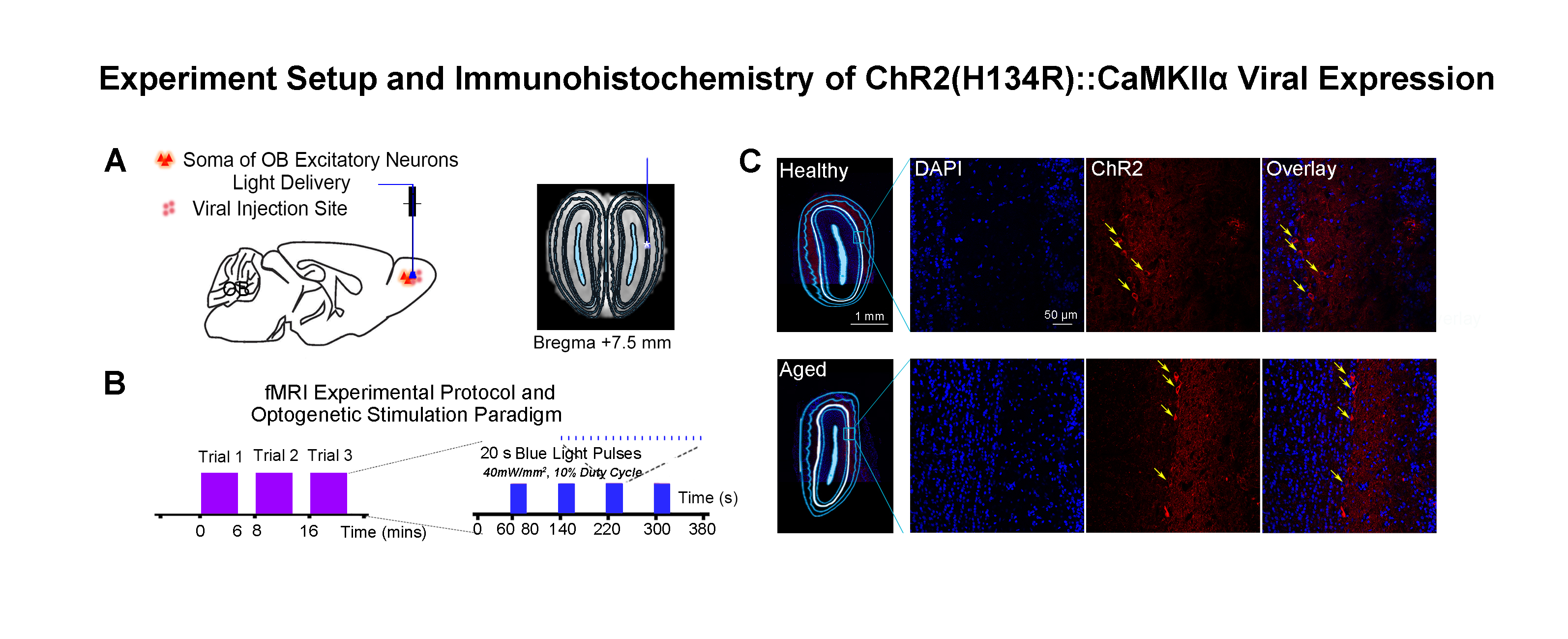
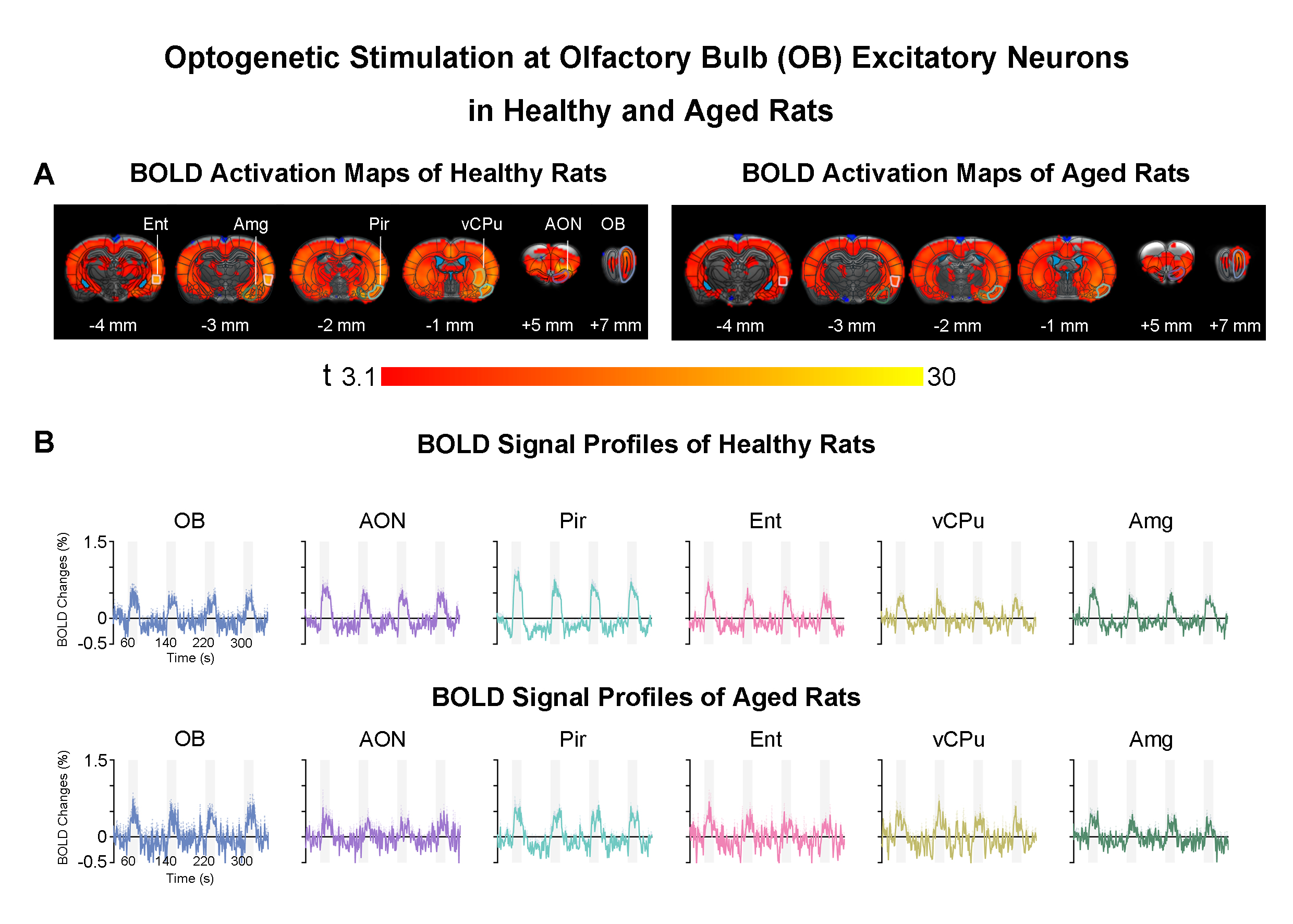
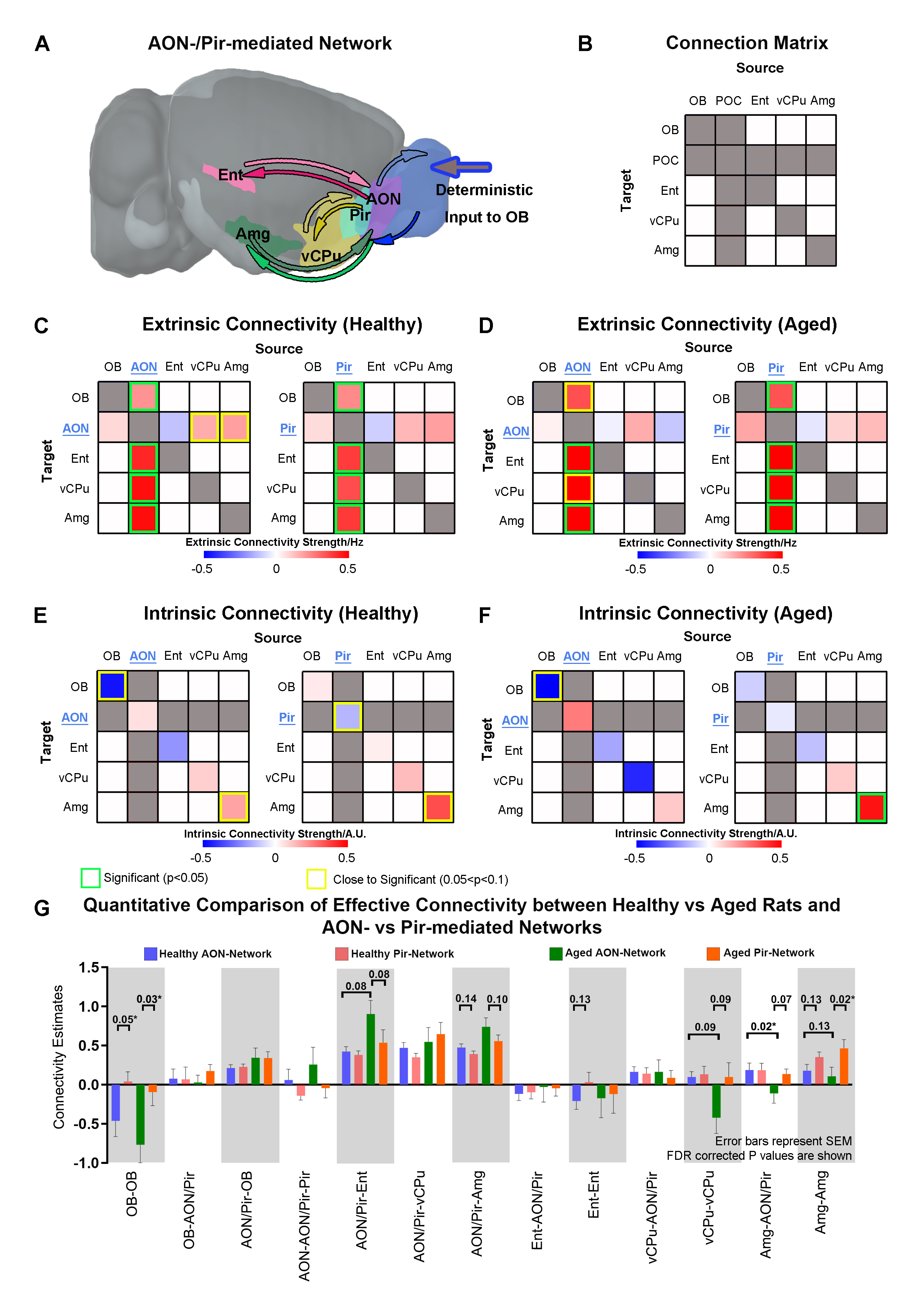
Accelerated aging rat model, optogenetic stimulation and fMRI acquisition and analysis: Adult rats (male, SD strain, 10 weeks old, n=8) were injected with D-galactose (100mg/kg) for 8 weeks3,4. At week five, 3μl of AAV5-CaMKIIα::ChR2(H134R)-mCherry was injected into OB (7.5mm anterior to Bregma, +1.7mm medial-lateral right hemisphere, -2.2mm from the surface of dura). Four weeks after viral injection, an opaque optical fiber cannula (d=450μm) was implanted at the injection site (Figure 1A). FMRI data was acquired on 7T Bruker scanner using GE-EPI (0.5x0.5x1mm3, α=56°, TE/TR=20/1000ms, and 20 contiguous slices). Data were preprocessed before standard GLM analysis was applied to identify significant BOLD responses (p<0.05; FDR corrected; Figure 2).DCM analysis5,6: Regions of interest (ROIs) were defined anatomically7-9 and the voxel-averaged fMRI time series were extracted. The priori connection schemes of anterior olfactory nucleus, AON-specific and piriform cortex, Pir-specific networks were defined (Figure 3A). After low-pass filtering the time series at 0.25 Hz, connectivity parameters were estimated for each subject by deterministic spectral DCM and significance of each connection was tested across subjects (healthy rats: n=10 and aged rats: n=8, t-test, p<0.05, FDR corrected). Regarding the optogenetic stimulation at OB as a single deterministic perturbation of the brain, observation noise was parameterized by different amplitude and autoregressive model coefficients (autoregressive models of order one, AR(1) to order four, AR(4))10.
Results
AON- and Pir-mediated networks reveal the diversity of effective connectivity (EC) in healthy and aged animals: Upon OB stimulation, significant extrinsic ECs were consistently observed across healthy and aged rats within OB-driven AON-/Pir-mediated networks, i.e., the feedforward ECs from AON/Pir to Ent/vCPu/Amg and feedback ECs from AON/Pir to OB, corroborating direct projection pathways11-13 (Figure 3C, D). We observed that only healthy animals displayed excitatory feedback EC from vCPu/Amg to AON. Meanwhile, AON-mediated OB self-disinhibition (negative intrinsic EC) and Pir-mediated Amg self-inhibition (positive intrinsic EC) were exhibited across healthy and aged animals, whereas AON-mediated Amg self-inhibition and Pir-mediated Pir self-inhibition were only exhibited in healthy, but not aged rats (Figure 3E, F).Furthermore, we quantitatively compared EC between healthy vs aged rats and AON-mediated vs Pir-mediated networks. An obvious increase trend of excitatory feedforward EC from AON to Ent was observed when comparing aged vs healthy rats. For intrinsic EC differences between healthy and aged animals, we found that vCPu & Amg self-disinhibition mediated by AON showed an increased trend across two groups (Figure 3G). It is notable that aging not only resulted in EC changes but also contributed to the EC diversity between AON- & Pir-networks.
Discussion & Conclusions
In this study, we examined the network-specific causal interactions in healthy and accelerated aging rat model using optogenetic fMRI and DCM. At present, there is no true consensus on the rat model of aging that best mimics normal aging processes in humans. D-galactose has been shown to induce advanced glycation end products formation, which contributes to early phases of age-related diseases in both humans and rodents14,15. Notably, not all olfactory-related functions will decline during aging. The preserved odor habituation1 and even increased emotion-associated memory in aging have been indicated in humans16 considering the age-dependent altered brain oscillations17. The increased trend in AON-mediated excitatory EC to limbic regions (i.e., Ent, Amg) and the decreased AON-mediated vCPu & Amg self-inhibition shown in our results corroborated this phenomenon. This may indicate the compensatory action of neural plasticity in which neural interactions are facilitated in aged brain to maintain olfactory function18,19. Further studies will implement stochastic DCM to account for endogenous fluctuations that cannot only be explained strictly by the experimental stimulus.Acknowledgements
This work was supported in part by Hong Kong Research Grant Council (HKU17112120, HKU17127121, HKU17127022 and HKU17127523 to E.X.W; and HKU17104020, HKU17127021, HKU17127723 to A.T.L.L.), Lam Woo Foundation, and Guangdong Key Technologies for AD Diagnostic and Treatment of Brain (2018B030336001) to E.X.W.References
1. Doty, R.L. Olfactory dysfunction in neurodegenerative diseases: is there a common pathological substrate? The Lancet Neurology 16, 478-488 (2017).
2. Mobley, A.S., Rodriguez-Gil, D.J., Imamura, F. & Greer, C.A. Aging in the olfactory system. Trends Neurosci 37, 77-84 (2014).
3. Ali, T., Badshah, H., Kim, T.H. & Kim, M.O. Melatonin attenuates D-galactose-induced memory impairment, neuroinflammation and neurodegeneration via RAGE/NF-K B/JNK signaling pathway in aging mouse model. J Pineal Res 58, 71-85 (2015).
4. Kay, L.M. Olfactory system oscillations across phyla. Curr Opin Neurobiol 31, 141-147 (2015).
5. Schott, B.H., Soch, J., Kizilirmak, J.M., Schutze, H., Assmann, A., Maass, A., Ziegler, G., Sauvage, M. & Richter, A. Inhibitory temporo-parietal effective connectivity is associated with explicit memory performance in older adults. iScience 26, 107765 (2023).
6. Zeidman, P., Jafarian, A., Seghier, M.L., Litvak, V., Cagnan, H., Price, C.J. & Friston, K.J. A guide to group effective connectivity analysis, part 2: Second level analysis with PEB. Neuroimage 200, 12-25 (2019).
7. Wang, X., Leong, A.T.L., Tan, S.Z.K., Wong, E.C., Liu, Y., Lim, L.W. & Wu, E.X. Functional MRI reveals brain-wide actions of thalamically-initiated oscillatory activities on associative memory consolidation. Nat Commun 14, 2195 (2023).
8. Leong, A.T.L., Gu, Y., Chan, Y.S., Zheng, H., Dong, C.M., Chan, R.W., Wang, X., Liu, Y., Tan, L.H. & Wu, E.X. Optogenetic fMRI interrogation of brain-wide central vestibular pathways. Proc Natl Acad Sci U S A 116, 10122-10129 (2019).
9. Leong, A.T., Chan, R.W., Gao, P.P., Chan, Y.S., Tsia, K.K., Yung, W.H. & Wu, E.X. Long-range projections coordinate distributed brain-wide neural activity with a specific spatiotemporal profile. Proc Natl Acad Sci U S A 113, E8306-E8315 (2016).
10. Bernal-Casas, D., Lee, H.J., Weitz, A.J. & Lee, J.H. Studying Brain Circuit Function with Dynamic Causal Modeling for Optogenetic fMRI. Neuron 93, 522-532 e525 (2017).
11. Gottfried, J.A. Central mechanisms of odour object perception. Nat Rev Neurosci 11, 628-641 (2010).
12. Albrecht, J. & Wiesmann, M. Olfactory Pathways. in Encyclopedia of Neuroscience (eds. Binder, M.D., Hirokawa, N. & Windhorst, U.) 3003-3006 (Springer Berlin Heidelberg, Berlin, Heidelberg, 2009).
13. Miyamichi, K., Amat, F., Moussavi, F., Wang, C., Wickersham, I., Wall, N.R., Taniguchi, H., Tasic, B., Huang, Z.J., He, Z.G., Callaway, E.M., Horowitz, M.A. & Luo, L.Q. Cortical representations of olfactory input by trans-synaptic tracing. Nature 472, 191-196 (2011).
14. Shen, Y.X., Xu, S.Y., Wei, W., Sun, X.X., Yang, J., Liu, L.H. & Dong, C. Melatonin reduces memory changes and neural oxidative damage in mice treated with D-galactose. J Pineal Res 32, 173-178 (2002).
15. Tian, J., Ishibashi, K., Ishibashi, K., Reiser, K., Grebe, R., Biswal, S., Gehlbach, P. & Handa, J.T. Advanced glycation endproduct-induced aging of the retinal pigment epithelium and choroid: A comprenensive transcriptionai response. Proceedings of the National Academy of Sciences of the United States of America 102, 11846-11851 (2005).
16. Jacques, P.S., Dolcos, F. & Cabeza, R. Effects of aging on functional connectivity of the amygdala during negative evaluation: A network analysis of fMRI data. Neurobiology of Aging 31, 315-327 (2010).
17. Ahnaou, A., Rodriguez-Manrique, D., Embrechts, S., Biermans, R., Manyakov, N.V., Youssef, S.A. & Drinkenburg, W.H.I.M. Aging Alters Olfactory Bulb Network Oscillations and Connectivity: Relevance for Aging-Related Neurodegeneration Studies. Neural Plasticity 2020(2020).
18. Morrissey, Z.D., Gao, J., Zhan, L., Li, W., Fortel, I., Saido, T., Saito, T., Bakker, A., Mackin, S., Ajilore, O., Lazarov, O. & Leow, A.D. Hippocampal functional connectivity across age in an App knock-in mouse model of Alzheimer's disease. Front Aging Neurosci 14, 1085989 (2022).
19. Zerbi, V., Wiesmann, M., Emmerzaal, T.L., Jansen, D., Van Beek, M., Mutsaers, M.P., Beckmann, C.F., Heerschap, A. & Kiliaan, A.J. Resting-state functional connectivity changes in aging apoE4 and apoE-KO mice. J Neurosci 34, 13963-13975 (2014).
Figures