0939
A deep dive into hippocampus growth: Unveiling neurite and soma development with in vivo diffusion MRI1Robarts Research Institute, Western University, London, ON, Canada, 2Department of Neurosurgery, The Royal Children's Hospital, Melbourne, Australia, 3Department of Radiology, New York University Grossman School of Medicine, New York, NY, United States, 4Cardiff University Brain Research Imaging Centre (CUBRIC), Cardiff University, Cardiff, United Kingdom, 5Department of Medical Biophysics, Western University, London, ON, Canada
Synopsis
Keywords: Microstructure, Aging, Hippocampus; High-field MRI
Motivation: The hippocampus serves multiple cognitive functions, yet little is known about its microstructural development.
Goal(s): To leverage recent MRI hardware and microstructure modeling advances to capture hippocampal cell-body (soma) and projection (neurite) development during late-childhood and adolescence.
Approach: Diffusion MRI data was acquired in 88 participants aged 8-18 years using a 3T Connectom scanner (with 300mT/m gradients), and analyzed using the Soma and Neurite Density Imaging model.
Results: For the first time, we identified distinct developmental patterns of hippocampal microstructural subcomponents. Specifically, we found an age-related increase in neurite fraction and concurrent decrease in extracellular fraction and soma radius.
Impact: We report, for the first time, distinct neurite and soma developmental profiles in the hippocampus during late childhood/adolescence. This forms a crucial baseline for understanding developmental disorders, and opens new avenues for corroborating in vivo diffusion with histology.
Introduction
The hippocampus is a widely studied yet enigmatic cortical region, and is classically divided into subfields based on cytoarchitecture or function1. Research using structural MRI has suggested that the hippocampal subfields undergo volume changes in childhood and adolescence, representing a likely expansion of cognitive capacity2. However, less is known about the development of the glial cells, neurites, and soma that form the microstructure of the hippocampus.The development of gray matter is characterized by microstructural changes including neurite (projection) growth, increased myelination, and a proliferation and subsequent pruning of synaptic connections3. Previous studies using diffusion tensor imaging4 to capture microstructural development of the hippocampus have found decreased diffusivity across childhood5. However, this finding is not specific to any particular microstructural property. Recent MRI advances including stronger gradients6 and new modeling approaches7 appear promising to disentangle soma and neurite contributions to the diffusion MRI signal in vivo.
Here we examined age-related changes of hippocampal microstructure using diffusion MRI (dMRI) data acquired with a 300 mT/m gradient in a sample of children and adolescents.
Methods
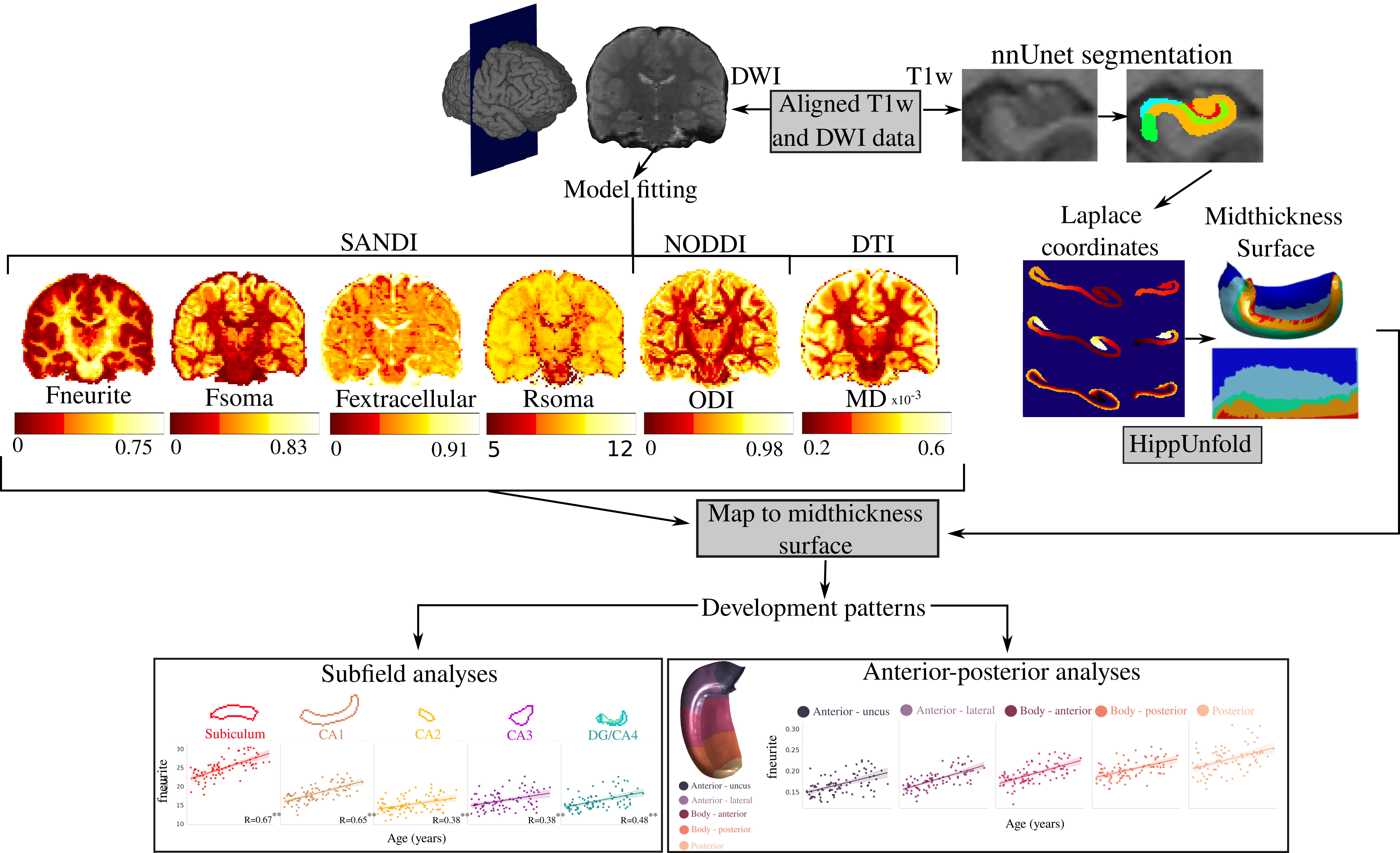
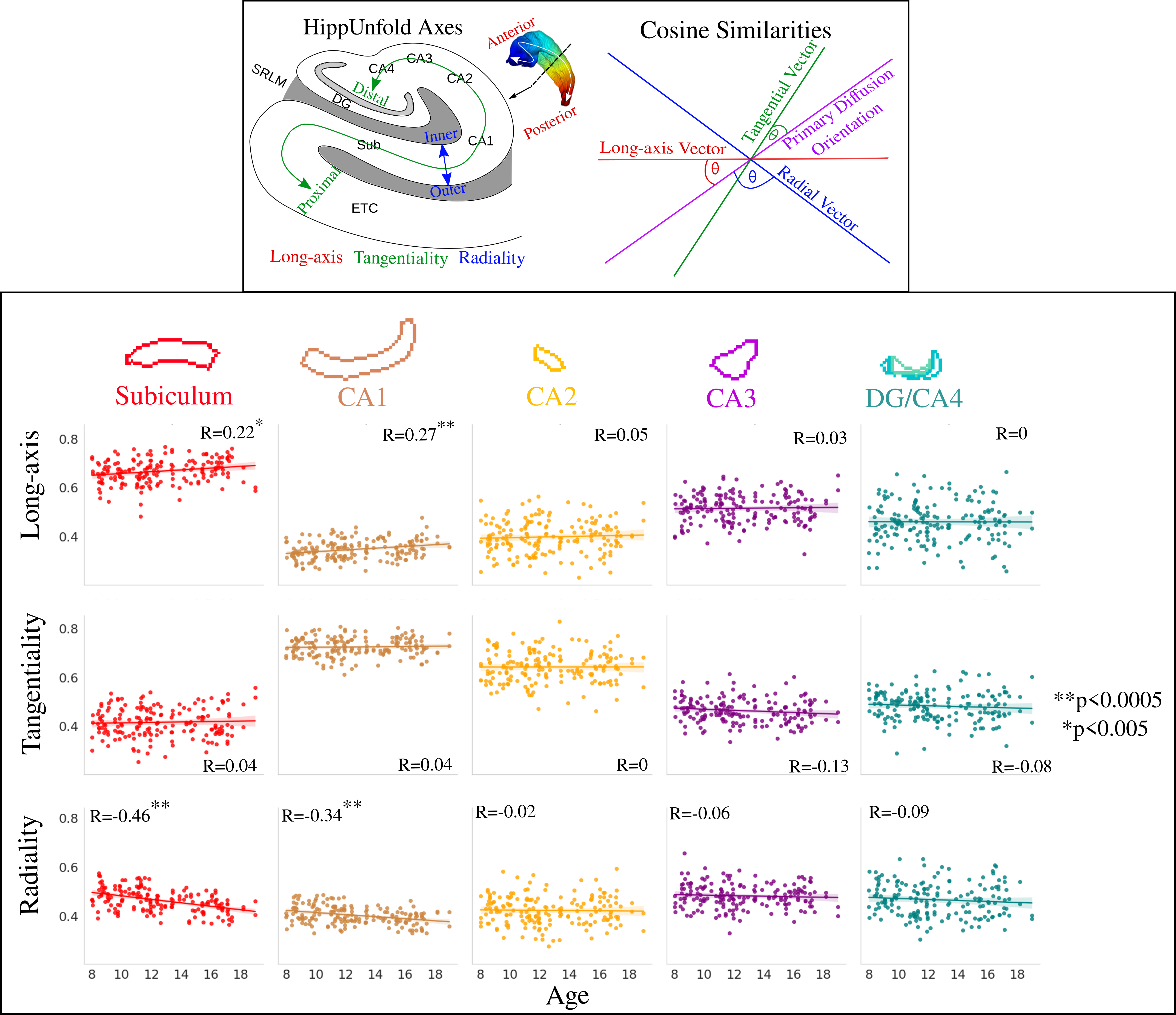
88 participants aged 8-18 years (42 male, mean age=12.6, SD=2.9) were scanned on a 3T Siemens Connectom system with 300 mT/m gradients8. T1-weighted images (1mm3) and dMRI (2mm3; b-values (directions)=0 (14), 500 (30), 1200 (30), 2400 (60), 4000 (60), 6000 (60) s/mm2) data were acquired8,9. HippUnfold10, was used for surface-based hippocampal modeling (Figure 1). The soma and neurite density imaging (SANDI)7 model was fitted to provide metrics of soma (fsoma), neurite (fneurite), and extracellular (fextracellular) signal fractions, and soma radius (Rsoma, in um). The neurite orientation dispersion and density imaging (NODDI)11 model was fitted to estimate the intracellular fraction (Vic) and orientation dispersion index (ODI). Finally, the DTI model was fitted using b-values of 0 and 6000 s/mm2 to derive mean diffusivity (MD in mm2/s), where higher b-values may increase sensitivity to cortical microstructure12. To interrogate diffusion orientations, cosine similarities were calculated as the dot product between hippocampal axis vectors (radial, tangential, and long-axis) and the primary diffusion orientation (Figure 5)13. All metrics were sampled on the midthickness surface from HippUnfold (Figure 1).Linear regression was performed across the subfields and long-axis to test for the effects of age, sex, and their interactions.
Results
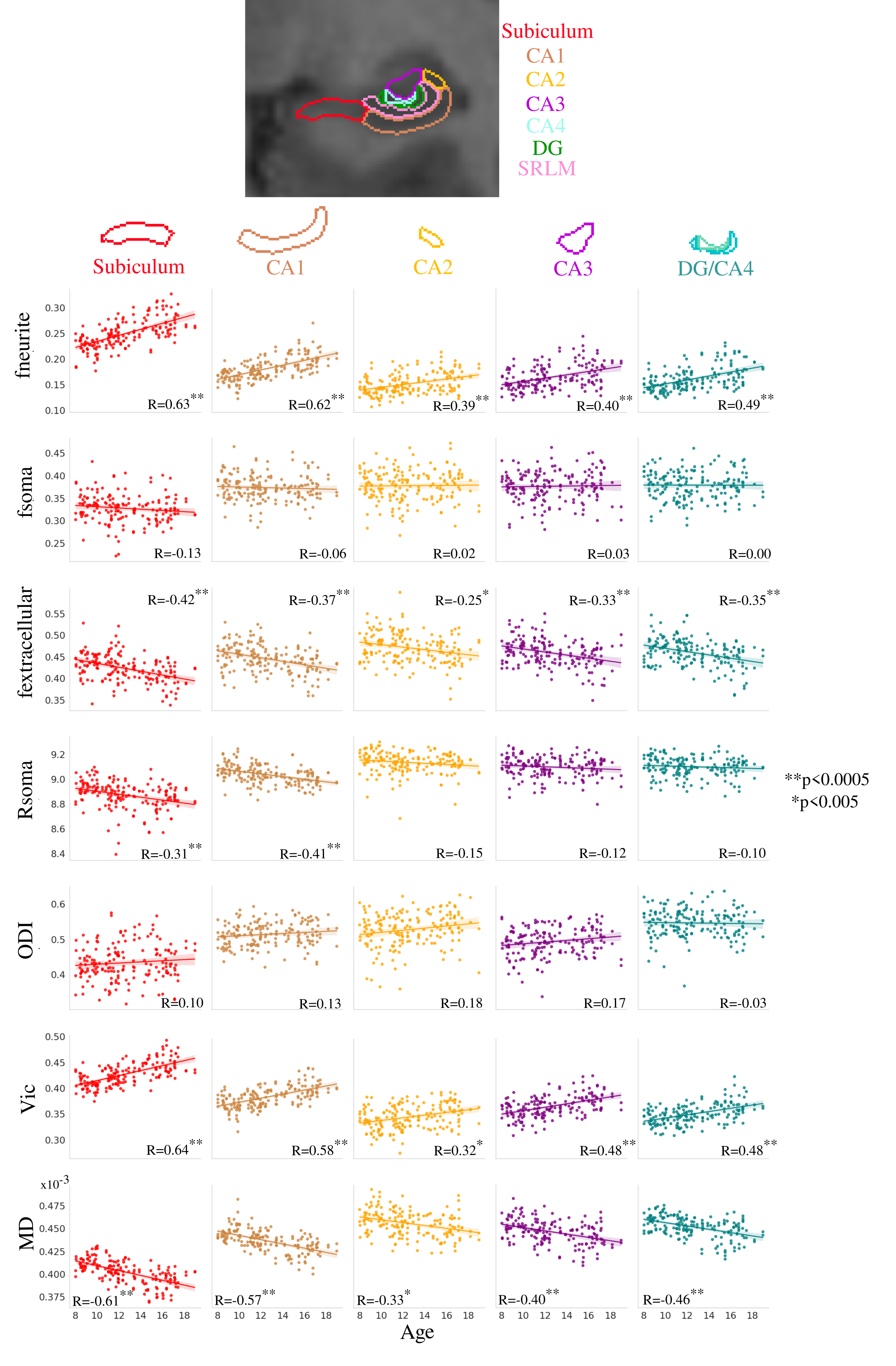
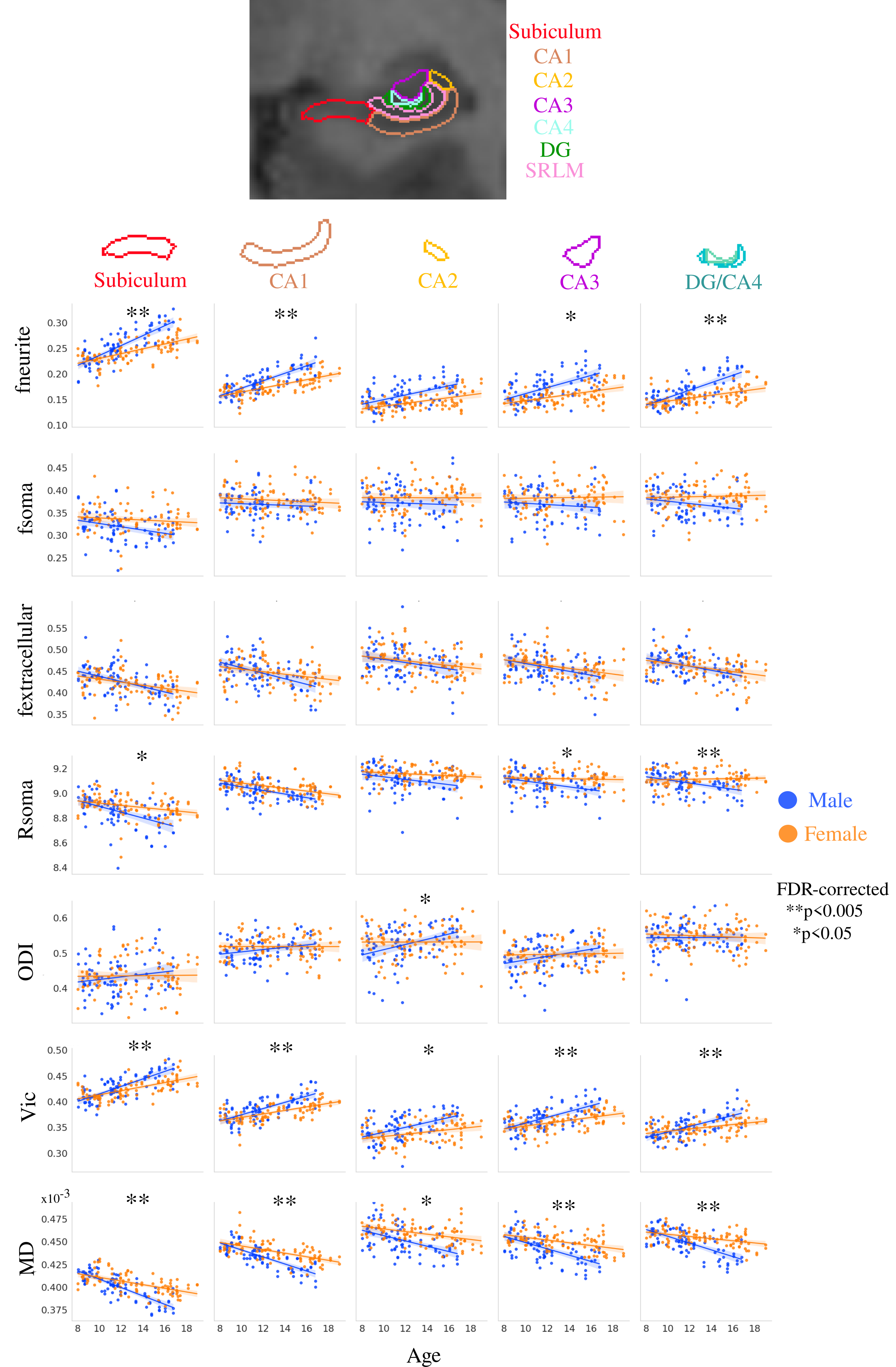
Distinct age-related microstructural patterns were observed within subfields for fneurite (mean R=0.51), fextracellular (mean R=-0.35), Rsoma in the subiculum and CA1 (R=-0.31, -0.41, respectively), Vic (mean R=0.50), and MD (mean R=-0.47), which is in-line with observations in the cortex8 (Figure 2). Furthermore the age-related changes were significantly different across subfields for fneurite (p<0.0005), Rsoma (p<0.005), and Vic (p<0.05), suggesting that different subfields have unique patterns of neurite and soma radius development.Significant within-subfield sex differences were found for development patterns of fneurite (males>females), fsoma (females>males), vic (males>females), and MD (females>males) (Figure 3).
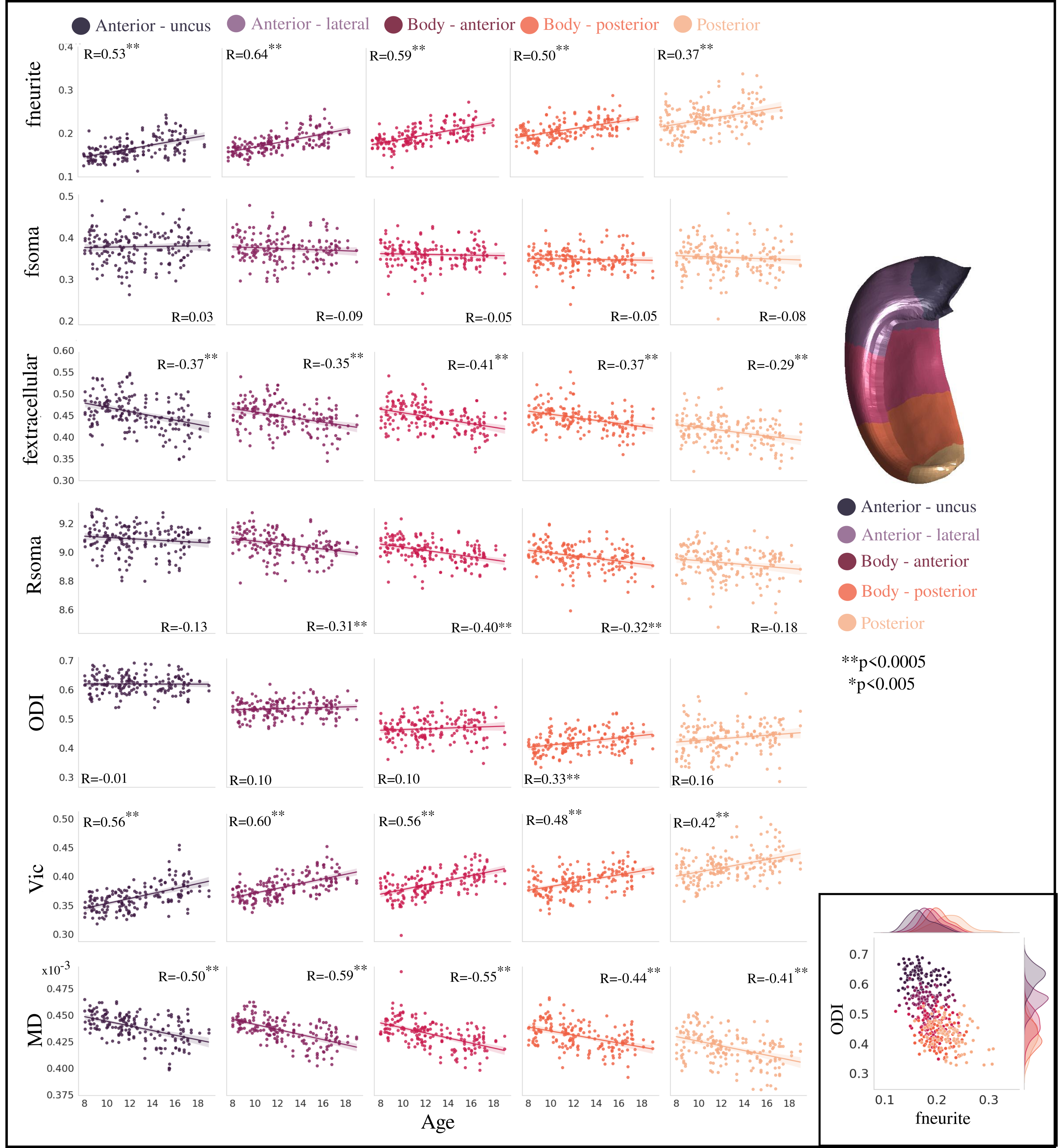
Examining microstructural development across the hippocampal long-axis, fneurite, vic, and posterior ODI were significantly higher with increasing age (Figure 4). Metrics of fextracellular, anterior and body Rsoma, and MD were significantly lower with increasing age. However, only ODI had different (p<0.05) age-related changes across the long-axis levels. The microstructural metrics themselves appear to vary across the long-axis (Figure 4 inset), suggesting that its microstructural environment is distinct.
A significant increase in long-axis oriented diffusion and significant decrease in radial diffusion in the subiculum and CA1 was observed with increasing age (Figure 5).
Discussion
Through a combination of novel diffusion MRI data and microstructural modelling, we have uncovered distinct developmental patterns in hippocampal microstructural sub-components. Subfield and long-axis neurite fraction increased significantly with age, potentially reflecting increased myelination14 and increased dendrite/astrocyte branching15. Age-related patterns differed across subfields for neurite fraction and soma radius, supporting the idea that the subfields have unique microstructural developmental patterns16. As well, no significant age-effects on soma fraction were found, partly in-line with an ex vivo study that found no difference in neuron number across development in any subfield except for the DG17.Radial diffusivity was also lower with age in the subiculum and CA1, which may be attributable to increased myelination of the perforant path in adolescence18, which itself is largely orthogonal to the hippocampal radial neurites.
Overall, our study uncovers unique patterns of hippocampal neurite and soma development in vivo. Future work should compare these results with histology to validate the age-dependent spatial patterns we report here.
Acknowledgements
The authors are grateful to the participants and their families for their participation in this study. We would like to thank Umesh Rudrapatna and John Evans for their support with MR acquisition, and Greg Parker and Chantal Tax for contributions to data preprocessing and model fitting pipelines.References
Voss JL, Bridge DJ, Cohen NJ, Walker JA. A closer look at the hippocampus and memory. Trends Cogn. Sci. 2017;21(8):577–88.
Tamnes CK, Bos MGN, van de Kamp FC, et al. Longitudinal development of hippocampal subregions from childhood to adulthood. Dev. Cogn. Neurosci. 2018;30:212–22.
Goldman-Rakic PS. Development of cortical circuitry and cognitive function. Child Dev.1987;58(3):601.
Basser PJ, Mattiello J, LeBihan D. MR diffusion tensor spectroscopy and imaging. Biophys. J. 1994;66(1):259–67.
Callow DD, Canada KL, Riggins T. Microstructural integrity of the hippocampus during childhood: Relations with age and source memory. Front. Psychol. 2020;11.
Jones DK, Alexander DC, Bowtell R, et al. Microstructural imaging of the human brain with a ‘super-scanner’: 10 key advantages of ultra-strong gradients for diffusion MRI. NeuroImage. 2018;182:8–38.
Palombo M, Ianus A, Guerreri M, et al. SANDI: A compartment-based model for non-invasive apparent Soma and neurite imaging by Diffusion MRI. NeuroImage. 2020;215:116835.
Genc S, Chamberland M, Ball G, et al. Disentangling neurite and soma contributions to developmental patterns of cortical microstructure in vivo. ISMRM Annual Meeting. 2021.
Genc S, Tax CM, Raven EP, et al. Impact of b‐value on estimates of apparent fibre density. Hum. Brain. Mapp. 2020;41(10):2583–95.
DeKraker J, Haast RA, Yousif MD, et al. Automated hippocampal unfolding for morphometry and subfield segmentation with Hippunfold. eLife. 2022;11.
Zhang H, Schneider T, Wheeler-Kingshott CA, Alexander DC. NODDI: Practical in vivo neurite orientation dispersion and density imaging of the human brain. NeuroImage. 2012;61(4):1000–16.
Mournet S, Okubo G, Koubiyr I, Zhang B, Kusahara H, Prevost VH, et al. Higher B-values improve the correlation between diffusion MRI and the cortical microarchitecture. Neuroradiology. 2020;62(11):1411–9. doi:10.1007/s00234-020-02462-4
Karat BG, DeKraker J, Hussain U, et al. Mapping the macrostructure and microstructure of the in vivo human hippocampus using diffusion MRI. Hum. Brain. Mapp. 2023;44(16):5485–503.
Ábrahám H, Vincze A, Jewgenow I, et al. Myelination in the human hippocampal formation from Midgestation to adulthood. Int. J. Dev. Neurosci. 2010;28(5):401–10.
Robillard KN, Lee KM, Chiu KB, MacLean AG. Glial cell morphological and density changes through the lifespan of rhesus macaques. Brain Behav Immun. 2016;55:60–9.
Wolf D, Fischer FU, de Flores R, et al. Differential Associations of age with volume and microstructure of hippocampal subfields in healthy older adults. Hum Brain Mapp. 2015;36(10):3819–31.
Jabès A, Lavenex PB, Amaral DG, Lavenex P. Postnatal development of the Hippocampal Formation: A stereological study in Macaque Monkeys. J. Comp. Neurol. 2011;519(6):1051–70.
Benes FM. Myelination of cortical-hippocampal relays during late adolescence. Schizoph. Bull. 1989;15(4):585–93.
Figures