0891
Laminar specific fMRI response is mainly regulated by first synaptic input-driven or more synchronous activity1Center for Neuroscience Imaging Research (CNIR), Suwon, Korea, Republic of, 2Korea Brain Research Institute (KBRI), Daegu, Korea, Republic of, 3Department of Biomedical Engineering, Sungkyunkwan University, Suwon, Korea, Republic of
Synopsis
Keywords: Task/Intervention Based fMRI, fMRI (task based)
Motivation: While responses to feedforward inputs have been well-observed in layer-specific fMRI studies, our understanding of responses to feedback projections within the ongoing functional processing remains limited.
Goal(s): Our study aimed to investigate how synaptic onset and strength contribute to specificity of laminar fMRI responses.
Approach: We performed the ultrahigh resolution CBV-weighted laminar fMRI by modulating thalamocortical and corticocortical projections with stimulus onset asynchrony in mice.
Results: We observed that the laminar response is highly sensitive to the strength of synaptic inputs, shifting from early to later input sites with increased strength.
Impact: Laminar CBV responses are highly regulated by micro-vessels coupled with earlier synaptic input activity, but potentially driven by the most synchronous activity within neural circuits.
Purpose
Recent advances in fMRI techniques at ultrahigh field have enabled the study of layer-specific activity, providing insights into information flow1-4. Laminar fMRI responses are suggested to mostly reflect neuronal input-driven activity, rather than output activity2. Functional networks contain multiple interconnected brain regions in a hierarchical and reciprocal manner. While responses to feedforward inputs have been well-observed (e.g., thalamus->S1->M1) in most layer-specific fMRI studies1-4, our understanding of responses to feedback projections within the ongoing functional processing (e.g., S1->M1->S1) remains limited. Therefore, we investigated whether laminar fMRI responses are primarily driven by first synaptic input layer and/or synaptic strength, achieved by modulating thalamocortical (TC) and corticocortical (CC) projections in mice.Materials & Methods
To separately replicate the patterns of somatosensory-evoked synaptic input for feedforward and feedback projections, we applied electrical whisker-pad stimulation (TC) and optogenetic excitation of whisker motor cortex (CC; AP: 1.0mm, ML: 1.2mm, and DV: 0.15mm) to transgenic Thy1-ChR2 mice (Fig.1A).All fMRI experiments were performed on 15.2 T MRI under medetomidine-isoflurane anesthesia1. To select a single slice containing S1BF responding to both whisker stimulation and optogenetic excitation, scout fMRI experiments were performed on the whole brain (Fig.1B). With two out-of-volume saturations (Fig.1C-i), CBV-weighted laminar fMRI data were acquired using gradient echo-based imaging (FLASH) with spatial resolution of 75x75x500um3 and temporal resolution of 2s (TE=3ms) following the injection of monocrystalline iron oxide nanoparticles (MION, 45 mg/kg) to enhance laminar specificity (Fig.1C-ii).
To ask the specificity of laminar fMRI response is determined by the contributions of feedforward vs. feedback inputs, we performed following experiments with 5 Hz stimulation (Fig. 2A). 1) TC inputs to S1BF by whisker-pad stimulation, 2) CC inputs to S1BF by optogenetic excitation of motor cortex (1mW), 3) TC-CC stimulus onset asynchrony with 50ms delay between two stimulus conditions (Fig.2A-iii), and 4) reversal of stimulus onset (Fig.2A-iv; n=8 mice). To enhance the strength of CC feedback inputs, experiment #2 and #4 were performed with 2 mW optogenetic excitation in a separate group (Fig.3A, n=8 mice).
To analyze the spatial distributions of fMRI responses, S1BF area was flattened by radially projecting 30 lines perpendicular to the cortical edges5,6 (Fig.2C). The cortical depth profiles were resampled to a nominal resolution of 50µm, and laminar boundaries were defined using cortical thickness distribution from the Allen mouse brain atlas.
Results
Animal-wise averaged fMRI maps and cortical depth profiles show that CBV-weighted responses were highly localized within the S1BF layers, corresponding to the neuronal input sites of TC (middle layer; Fig.2B-i, D-i and Fig.3B-i, D-i) and CC (upper layer; Fig.2B-ii, D-ii and Fig.3B-ii, D-ii) projections, which is consistent with our previous fMRI studies4,6. Intriguingly, even when whisker stimulation preceded optogenetic excitation of the motor cortex, the highest fMRI responses still occurred in S1BF middle layer (Fig.2B-iii, D-iii and Fig.3B-iii, D-iii). Conversely, when the order of stimulus onset asynchrony was reversed, the response of CC projection induced by optogenetic excitation still remained predominant (Fig.2B-iv, D-iv). These indicates that the hemodynamic response is mostly sensitive to the first synaptic input layer. However, when the secondary input strength is larger than the first synaptic input strength, the second inputs also contribute to CBV response (Fig.3B-iv, D-iv).Discussion & Conclusion
We aimed to examine the contribution of both synaptic onset and strength to specificity of the laminar fMRI responses with TC and CC projections to S1BF. In our study, fMRI activations were predominantly observed in layers responsive to first synaptic inputs. This is consistent with previous findings from electrophysiological recordings7 and fMRI experiments8. The second synaptic inputs (50 ms delay) may be suppressed by the initial inputs, which is required to be confirmed by electrophysiology in our experimental setup. When the strength of the secondary stimulus train increased, the fMRI peak shifted from layers for the initial inputs to active sites corresponding to the later inputs. This suggests that laminar CBV responses are highly regulated by micro-vessels coupled with earlier synaptic input activity, but potentially driven by the most synchronous activity within neural circuits.Overall, hemodynamic responses are expected to increase with the magnitude of coherent input-driven activity9. In functional network, wherever neural projections occur, synaptic input activity at the downstream sites is likely to be less synchronized, causing a challenge in detecting secondary feedback inputs (e.g., S1->M1->S1).
Acknowledgements
This work was supported by IBS-R015-D1References
1. Lawrence SJD, Formisano E, Muckli L, de Lange FP. Laminar fMRI: Applications for cognitive neuroscience. Neuroimage. 2019;197:785-791.
2. Huber L, Handwerker DA, Jangraw DC, et al. High-Resolution CBV-fMRI Allows Mapping of Laminar Activity and Connectivity of Cortical Input and Output in Human M1. Neuron. 2017;96(6):1253-1263.e7. 3. Yang J, Huber L, Yu Y, Bandettini PA. Linking cortical circuit models to human cognition with laminar fMRI. Neurosci Biobehav Rev. 2021;128:467-478.
4. Jung WB, Jiang H, Lee S, Kim SG. Dissection of brain-wide resting-state and functional somatosensory circuits by fMRI with optogenetic silencing. Proc Natl Acad Sci U S A. 2022;119(4):e2113313119.
5. Shih YY, Chen YY, Lai HY, Kao YC, Shyu BC, Duong TQ. Ultra high-resolution fMRI and electrophysiology of the rat primary somatosensory cortex. Neuroimage. 2013;73:113-120.
6. Jung WB, Im GH, Jiang H, Kim SG. Early fMRI responses to somatosensory and optogenetic stimulation reflect neural information flow. Proc Natl Acad Sci U S A. 2021;118(11):e2023265118.
7. Lohse, M., Cooper, M., Sader, E., et al., Motor cortex can modulate somatosensory processing via cortico-thalamo-cortical pathway. bioRxiv, 2018;366567.
8. Chen Y, Sobczak F, Pais-Roldán P, Schwarz C, Koretsky AP, Yu X. Mapping the Brain-Wide Network Effects by Optogenetic Activation of the Corpus Callosum. Cereb Cortex. 2020;30(11):5885-5898
9. Logothetis NK, Pauls J, Augath M, Trinath T, Oeltermann A. Neurophysiological investigation of the basis of the fMRI signal. Nature. 2001;412(6843):150-157.
Figures
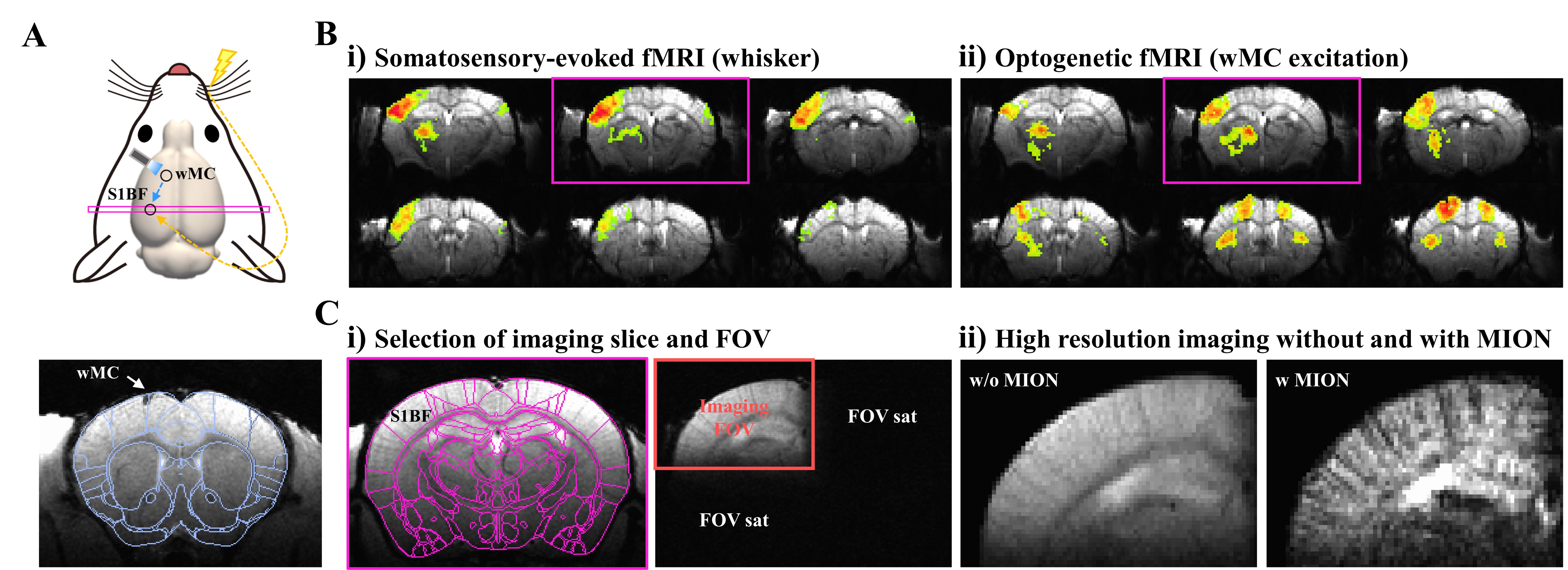
Figure 1. Experimental setup for ultrahigh resolution CBV-weighted laminar fMRI in mice
A. Schematic of experimental design to induce fMRI responses in S1BF by electrical whisker-pad stimulation and optogenetic excitation of motor cortex
B. Scout fMRI of the whole brain to select a slice covering the somatosensory cortex of barrel field
C. Selection of imaging FOV for ultrahigh resolution CBV-weighted laminar fMRI
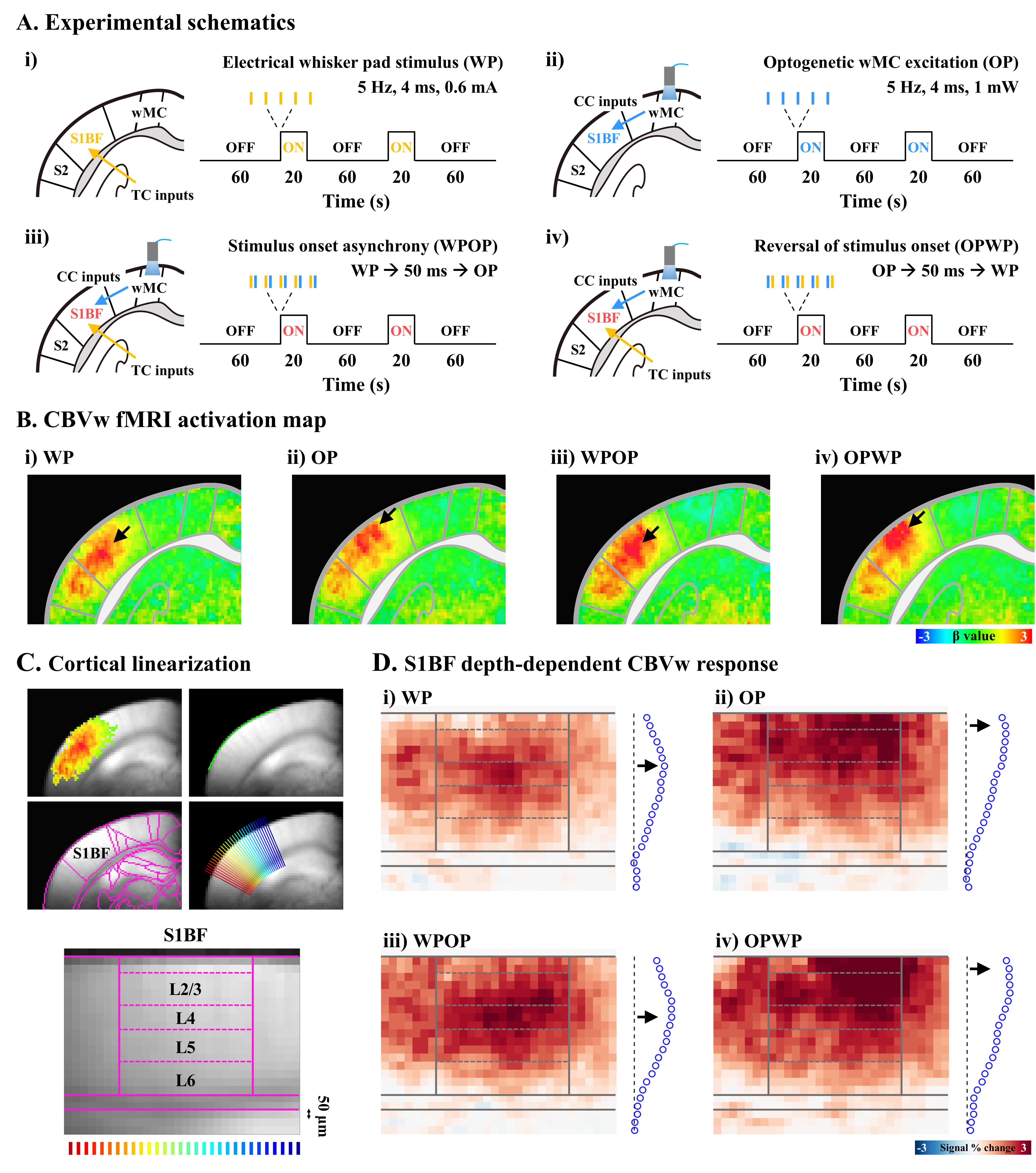
Figure 2. CBV-weighted response is sensitive to the neuronal active sites to the early input train.
A. Experimental schematics of four different stimulus conditions
B. Group-averaged fMRI activation maps
C. Cortial flattening for layer-specific fMRI analysis in the S1BF
D. Cortical depth-dependent fMRI maps
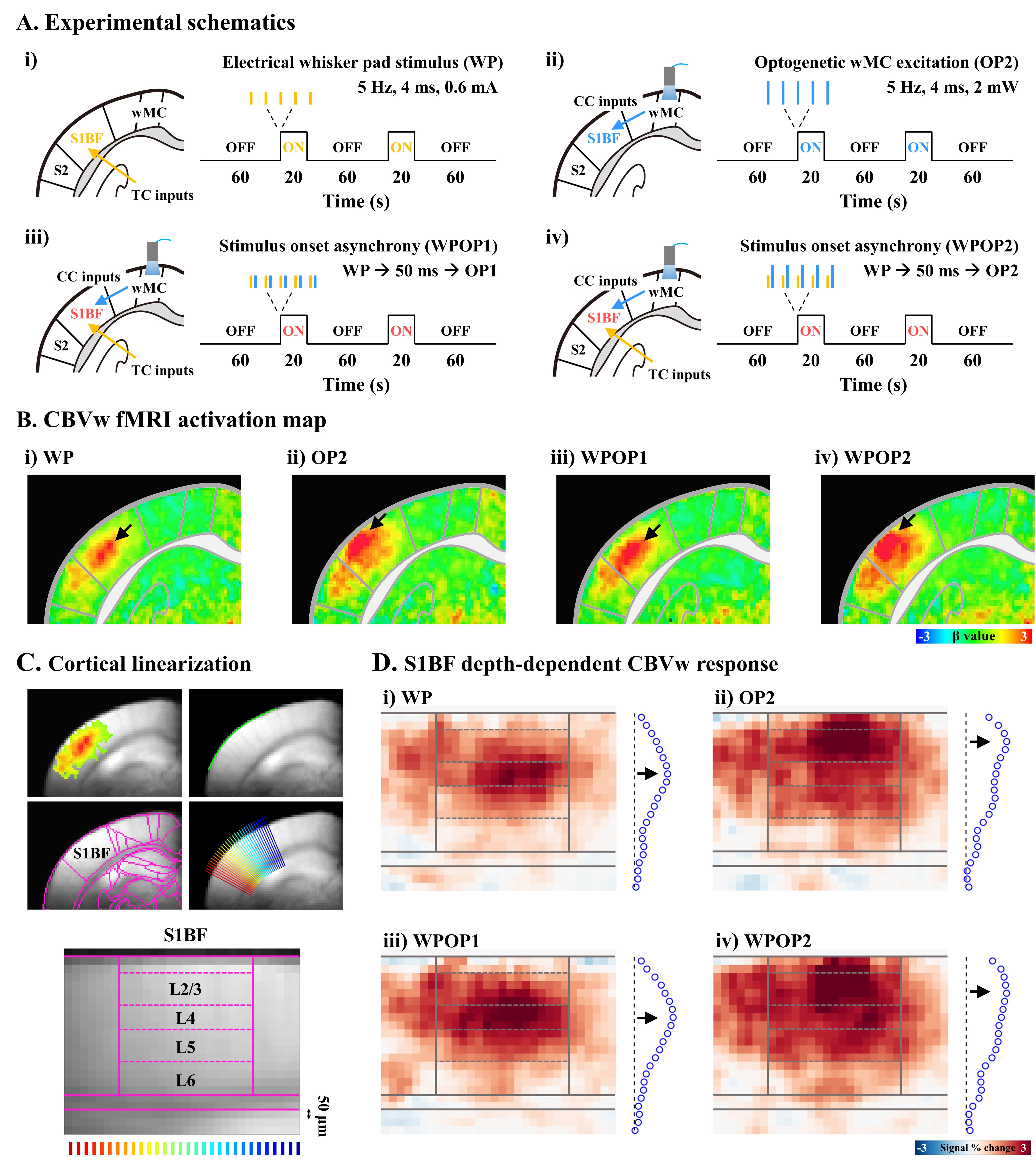
Figure 3. The highest CBV laminar response is driven by the most synchronous activity
A. Experimental schematics of four different stimulus conditions
B. Group-averaged fMRI activation maps
C. Cortial flattening for layer-specific fMRI analysis in the S1BF
D. Cortical depth-dependent fMRI maps