0884
Characterisation of cortical depth dependent hemodynamics in early human development using high-resolution BOLD fMRI at 7 Tesla.1Centre for the Developing Brain, School of Biomedical Engineering and Imaging Sciences, Kings College London, London, United Kingdom, 2MRC Centre for Neurodevelopmental Disorders, King's College London, London, United Kingdom, 3Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital, Charlestown, MA, United States, 4Guy's and St Thomas' NHS Foundation Trust, London, United Kingdom, 5London Collaborative Ultra high field System (LoCUS), King's College London, London, United Kingdom, 6Biomedical Engineering Department, School of Biomedical Engineering and Imaging Sciences, King's College London, London, United Kingdom, 7Department of Radiology, Harvard Medical School, Boston, MA, United States, 8Harvard-MIT Division of Health Sciences and Technology, Massachusetts Institute of Technology, Cambridge, MA, United States
Synopsis
Keywords: fMRI Analysis, High-Field MRI, Brain, Neuro
Motivation: Hemodynamic responses in adults vary across cortical depths, partly due to specific differences in vascular anatomy and physiology. It is unknown how these differences relate to the responses seen in neonates, when the cortex and neurovasculature are rapidly maturing.
Goal(s): To characterize the amplitude and timing of the hemodynamic response across cortical depths during the neonatal period.
Approach: Cortical depth-dependent hemodynamic responses to sensorimotor stimulation were delineated using GRE-BOLD fMRI in term-aged neonates.
Results: A cortical depth-specific profile of key hemodynamic response parameters, including onset and undershoot, was observed in the neonatal cortex that differs from those seen in the adult.
Impact: We demonstrate for the first time how developing vascular network may alter hemodynamic response across cortical depths. It illuminates what underlying neurobiology may generate the neonatal specific BOLD signal profile and what components are altered relative to the adult response.
Introduction
Across the perinatal period, neuronal activity, neurovascular coupling and vascular dynamics all rapidly mature1–6. As a result, in the newborn period, the hemodynamic response to neuronal activity differs markedly from that of adults, with slower time to peak, broader temporal dispersion, and decreased amplitude7,8. In adults, cortical depth-dependent differences in hemodynamic responses features–such as timing and presence of a post-stimulus BOLD undershoot–are observed, which likely reflect differences in hemodynamics across different levels of the intracortical vascular hierarchy9,10 and, potentially cortical depth-dependent differences in tissue stiffness and vascular compliance11,12. However, it is unclear whether this understanding can be generalised to the developing neonatal system as across development there is ongoing arterial maturation, increasing depth-specific parenchymal microvascular density, and altered post-capillary vascular compliance 2,4,6,7,13. To address this, we developed a platform for high-resolution fMRI in neonates at 7 Tesla and performed a cortical-depth analysis of cortical BOLD responses. Given known differences in vascular anatomy and physiology, we hypothesised that neonatal hemodynamic responses would comprise a different profile across cortical depths compared with that seen in the adult brain.Methods
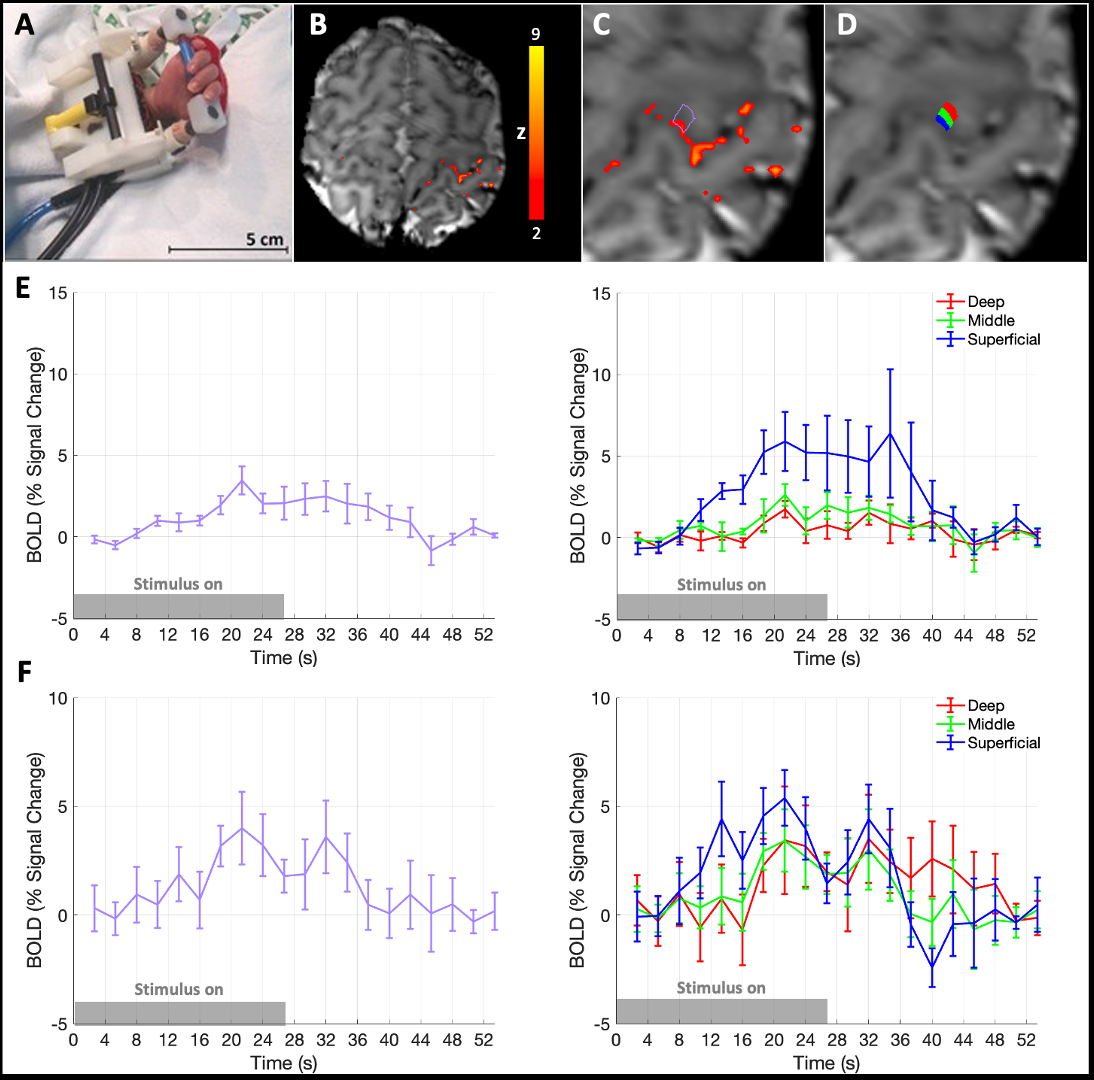
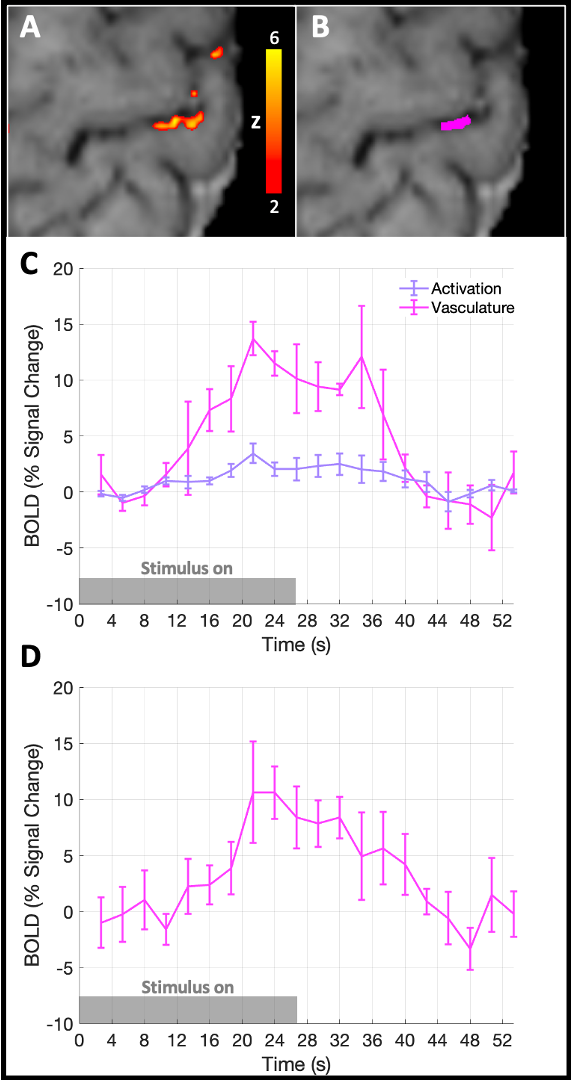
NHS research ethics committee approval and written parental consent was attained for data collection.Data were acquired from 10 healthy term-age neonates (median: 39.7 weeks post-menstrual age at scan, range: 37.4–42.9 weeks, 5 female) during natural sleep using a Siemens 7T system (MAGNETOM Terra, Siemens Healthineers, Erlangen, DE) and a 1TX-32RX Nova Medical head coil (Wilmington, MA, USA) with locally imposed safety restrictions14. BOLD-weighted fMRI data were acquired with a GRE-EPI sequence over 6 m 51 s with parameters: resolution=0.8 mm isotropic, TR/TE=2660/48 ms, 25 slices, 1.06 ms nominal echo spacing, R=2 acceleration with Dual-Polarity GRAPPA reconstruction15. A custom built robotic device provided sensorimotor stimulation (right 0.33 Hz wrist extension/flexions, in on/off blocks of 26.6 s)16. After standard pre-processing, whole cortical thickness ROIs were manually defined within significant clusters of activation (Z>2) in the primary motor cortex (M1) hand area; data were up-sampled to 0.16 x 0.16 x 0.8 mm3 resolution to avoid singularities at the cortical boundaries and 3 equi-volume layers were defined using LAYNII17. Layer specific BOLD timeseries were extracted and averaged. Venous ROIs were also defined in the same manner but constrained to clusters of activation with clear pial vasculature origin.
Results
fMRI data were successfully acquired 7/10 infants. Localised activation clusters were seen in the contralateral hand area of M1 in response to sensorimotor stimulation in 5/7 infants (Fig. 1A, B).The trial-averaged response showed a rise in BOLD signal around 8 s, followed by a post-stimulus undershoot that was most distinct in the superficial layers (Fig. 1E, F). Conversely, the BOLD response in deep layers comprised a small initial dip followed by a delayed rise in BOLD signal, a lower peak amplitude and reduced post-stimulus undershoot at the deep cortical layers. Activation was also localised to a distinct pial vein adjacent to a second cluster of activation in 4/7 infants. BOLD responses in these clusters showed a later onset, larger peak amplitude and more pronounced post-stimulus undershoot (Fig. 2).
Discussion
We demonstrate that neonatal hemodynamic responses have different properties across cortical depth compared with adults.Diving arterioles dilate first in the deeper cortical depths in response to neuronal activity, with vasodilation propagating upstream along arterioles to the pial surface18,19. In neonates, the vascular endothelium of arterioles develops postnatally, perhaps leading to maturational differences in the vasodilation propagation2 and delayed rise in BOLD response in the deeper cortical layers.
We observed a more pronounced BOLD post-stimulus undershoot superficially than at deeper layers, perhaps due to reduced elasticity and thus slower recoil of the post-capillary vasculature leading to venous “ballooning” and accumulation of deoxygenated hemoglobin near the pial surface7,20. Additionally, cortical grey matter stiffness increases across development as water content decreases and tissue density rises with neuronal arborisation21, and therefore passive compliance of veins may be greater in neonates11,12.
Although stimuli processing and neurovascular coupling are grossly maintained during sleep, we cannot exclude the possibility of sleep effects on the observed trends22,23.
Conclusion
The increased functional contrast-to-noise ratio at 7 Tesla24 provides a unique opportunity for detailed studies of neurovascular coupling and hemodynamics in vivo during the critical period around birth. Here, we present the first evidence of differing hemodynamic responses across cortical depths in the neonatal brain. This offers new insight into the early development of cortical hemodynamics and provides a model system to explore generalisability of the mechanistic understanding of adult cortical hemodynamics. Moreover, accounting for these differences will be important for interpreting laminar fMRI analyses in the early developing brain.Acknowledgements
This work was supported by a project grant awarded by Action Medical Research [GN2728], an MRC Clinician Scientist Fellowship [MR/P008712/1], an MRC Transition Support Award [MR/V036874/1], the KCL MRC Centre for Neurodevelopmental Disorders [MR/N026063/1], core funding from the Wellcome/EPSRC Centre for Medical Engineering [WT 203148/Z/16/Z], the National Institute for Health Research (NIHR) Clinical Research Facility based at Guy’s and St Thomas’ NHS Foundation Trust and King’s College London, and by the NIH NIBIB (grants P41-EB030006). We also thank Siemens Healthineers for providing the Works-In-Progress (WIP) package 1105D used in this research. The views expressed are those of the author(s) and not necessarily those of the NHS, the NIHR or the Department of Health and Social Care.References
1. Attwell, D. & Iadecola, C. The neural basis of functional brain imaging signals. Trends Neurosci. 25, 621–625 (2002).
2. Kozberg, M. & Hillman, E. Neurovascular coupling and energy metabolism in the developing brain. Prog. Brain Res. 225, 213–242 (2016).
3. Kozberg, M. G., Ma, Y., Shaik, M. A., Kim, S. H. & Hillman, E. M. C. Rapid Postnatal Expansion of Neural Networks Occurs in an Environment of Altered Neurovascular and Neurometabolic Coupling. J. Neurosci. 36, 6704–6717 (2016).
4. Norman, M. G. & O’Kusky, J. R. The Growth and Development of Microvasculature in Human Cerebral Cortex. J. Neuropathol. Exp. Neurol. 45, 222–232 (1986).
5. Harris, J. J., Reynell, C. & Attwell, D. The physiology of developmental changes in BOLD functional imaging signals. Dev. Cogn. Neurosci. 1, 199–216 (2011).
6. Coelho-Santos, V., Berthiaume, A.-A., Ornelas, S., Stuhlmann, H. & Shih, A. Y. Imaging the construction of capillary networks in the neonatal mouse brain. Proc. Natl. Acad. Sci. 118, e2100866118 (2021).
7. Arichi, T. et al. Development of BOLD signal hemodynamic responses in the human brain. Neuroimage 63, 663–673 (2012).
8. Kozberg, M. G., Chen, B. R., DeLeo, S. E., Bouchard, M. B. & Hillman, E. M. C. Resolving the transition from negative to positive blood oxygen level-dependent responses in the developing brain. Proc. Natl. Acad. Sci. 110, 4380–4385 (2013).
9. Siero, J. C. W. et al. BOLD Specificity and Dynamics Evaluated in Humans at 7 T: Comparing Gradient-Echo and Spin-Echo Hemodynamic Responses. PLoS ONE 8, e54560 (2013).
10. Siero, J. C. W. et al. Cortical depth dependence of the BOLD initial dip and poststimulus undershoot in human visual cortex at 7 Tesla. Magn. Reson. Med. 73, 2283–2295 (2015).
11. Pfannmoeller, J. P. et al. Simulations of the BOLD Non-Linearity Based on a Viscoelastic Model for Capillary and Vein Compliance. in Proceedings of the International Society for Magnetic Resonance in Medicine. vol. 29 2856 (2021).
12. Gao, Y.-R., Greene, S. E. & Drew, P. J. Mechanical restriction of intracortical vessel dilation by brain tissue sculpts the hemodynamic response. NeuroImage 115, 162–176 (2015).
13. Tieman, S. B., Möllers, S., Tieman, D. G. & White, J. The blood supply of the cat’s visual cortex and its postnatal development. Brain Res. 998, 100–112 (2004).
14. Malik, S. J., Hand, J. W., Satnarine, R., Price, A. N. & Hajnal, J. V. Specific absorption rate and temperature in neonate models resulting from exposure to a 7T head coil. Magn. Reson. Med. 86, 1299–1313 (2021).
15. Hoge, W. S. & Polimeni, J. R. Dual-Polarity GRAPPA for simultaneous reconstruction and ghost correction of EPI data. Magn. Reson. Med. 76, 32–44 (2016).
16. Allievi, A. G., Melendez-Calderon, A., Arichi, T., Edwards, A. D. & Burdet, E. An fMRI Compatible Wrist Robotic Interface to Study Brain Development in Neonates. Ann. Biomed. Eng. 41, 1181–1192 (2013).
17. Huber, L. (Renzo) et al. LayNii: A software suite for layer-fMRI. NeuroImage 237, 118091 (2021).
18. Tian, P. et al. Cortical depth-specific microvascular dilation underlies laminar differences in blood oxygenation level-dependent functional MRI signal. Proc. Natl. Acad. Sci. 107, 15246–15251 (2010).
19. Uhlirova, H. et al. Cell type specificity of neurovascular coupling in cerebral cortex. eLife 5, e14315 (2016).
20. Buxton, R. B., Uludağ, K., Dubowitz, D. J. & Liu, T. T. Modeling the hemodynamic response to brain activation. NeuroImage 23, S220–S233 (2004).
21. Schneider, J. et al. Evolution of T1 Relaxation, ADC, and Fractional Anisotropy during Early Brain Maturation: A Serial Imaging Study on Preterm Infants. Am. J. Neuroradiol. 37, 155–162 (2016).22. Portas, C. M. et al. Auditory Processing across the Sleep-Wake Cycle: Simultaneous EEG and fMRI Monitoring in Humans. Neuron 28, 991–999 (2000).
23. Turner, K. L., Gheres, K. W., Proctor, E. A. & Drew, P. J. Neurovascular coupling and bilateral connectivity during NREM and REM sleep. eLife 9, e62071 (2020).
24. Viessmann, O. & Polimeni, J. R. High-resolution fMRI at 7 Tesla: challenges, promises and recent developments for individual-focused fMRI studies. Curr. Opin. Behav. Sci. 40, 96–104 (2021).
Figures