0764
Sex Specific metabolism of Branched Chain Amino Acids and Relation with Cerebral Glutamate in Alzheimer’s Disease: A 1H-MRS Study1Department of Radiology, University of Pennsylvania, Philadelphia, PA, United States
Synopsis
Keywords: Alzheimer's Disease, Alzheimer's Disease, 1H MRS, BCAAs, Glutamate
Motivation: Amyloid-β and NFTs are considered hallmarks of Alzheimer’s disease (AD) though are often challenged. While branched chain amino acids (BCAA) are known to play role in AD pathogenesis, their role is underexplored. A non-invasive method to study this relation could benefit devising alternate strategies for early AD diagnosis.
Goal(s): To monitor sex-specific changes in BCAA levels and its relationship with glutamate.
Approach: Localized-1H-MRS was performed in AD and WT mice.
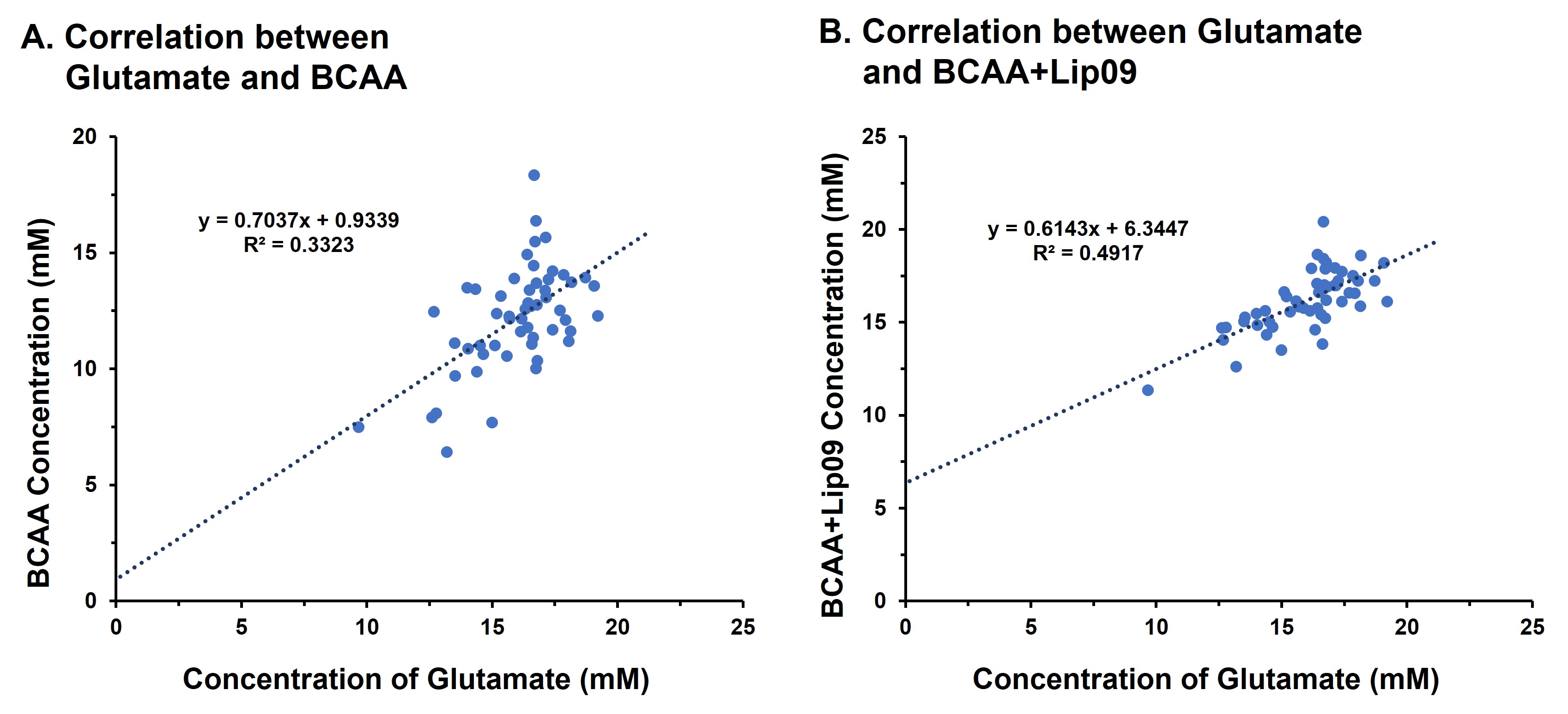
Results: Significant reduction in hippocampal BCAA and glutamate levels in male AD mice were observed, while females remained unaffected. BCAA and glutamate levels shown a strong correlation suggesting their close association in AD.
Impact: Establishing a relation between BCAA metabolism and AD pathology could be beneficial for preclinical diagnosis as clear resonances of BCAA around 0.9 ppm on 1H MR spectra is achievable even at lower field strength MRI scanners.
Introduction
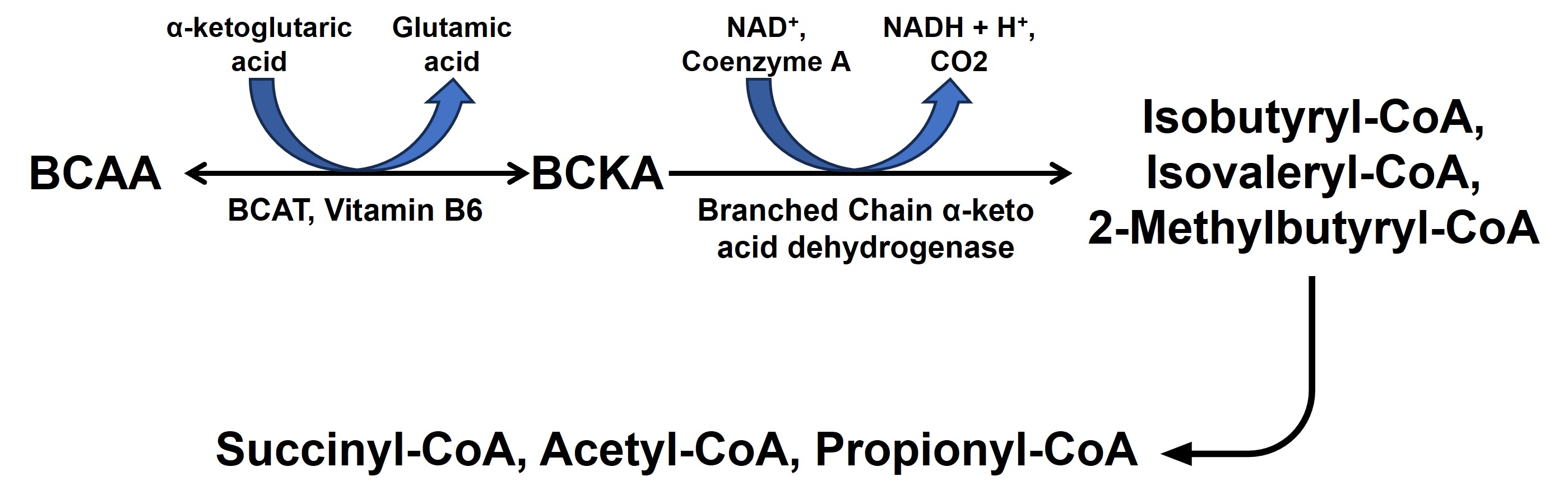
Alzheimer’s disease (AD) is the most common neurodegenerative disorder characterized by progressive loss of cognitive and intellectual abilities including memory1. Branched-chain amino acids (BCAAs) comprise of valine, leucine and isoleucine and studies have shown that altered BCAAs metabolism accompanies AD development. Lower plasma valine levels are correlated with accelerated cognitive decline, and, conversely, an increase in valine concentration is associated with a reduced risk of AD2. Glutamate, the major excitatory neurotransmitter in the mammalian central nervous system3 plays an important role in regulation of cognitive functions including learning and memory4. Multiple studies conducted on AD animal models5,6 and human subjects7 using 1H MRS have revealed a consistent reduction in the levels of cortical glutamate. Also, the metabolism of BCAA is coupled with the replenishment of glutamate and other TCA intermediates (Fig. 1). It will be very interesting to monitor the BCAA levels and their correlation with glutamate levels in the early stage of AD. In the current study, our objective was to perform 1H-MRS in the hippocampus of 6-month-old APPNL-F/NL-F male and female mice. The presence of a prominent sex bias in the severity, prevalence, and progression of AD drove us to conduct this study in a sex-specific manner.Methods
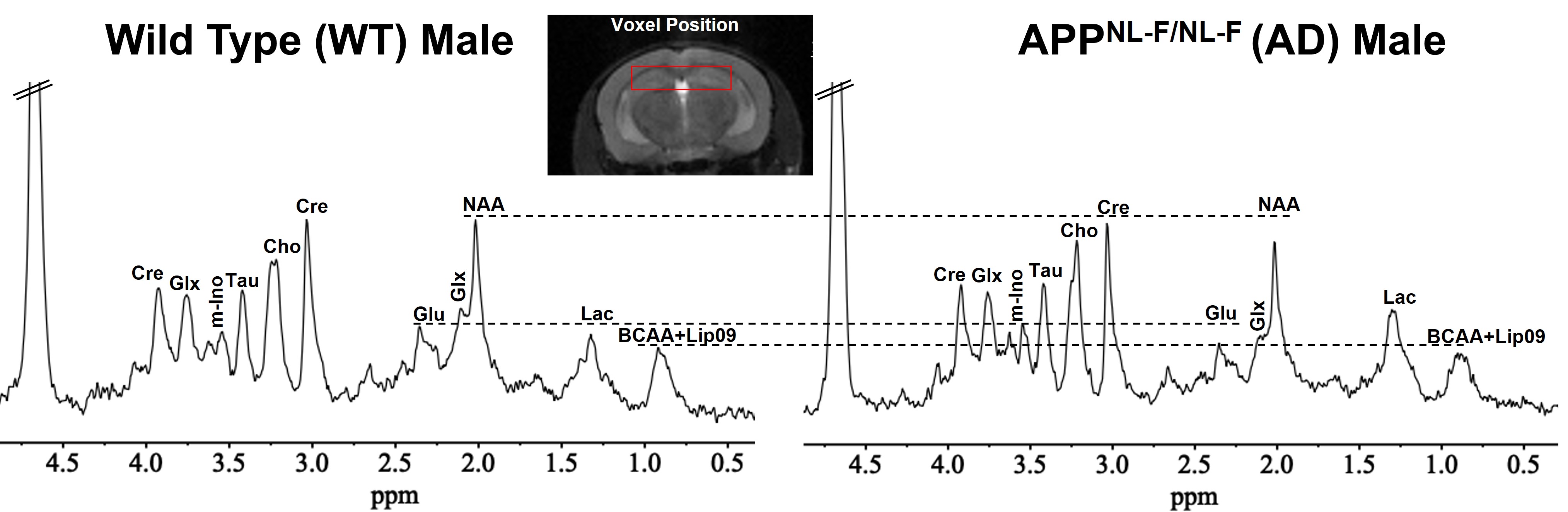
The protocol used for animal experiments was approved by the IACUC committee of the University of Pennsylvania. The study was performed using 6-month-old C57BL6/J (Wild Type (WT), n=24; 15 male and 9 female) and APPNL-F/NL-F (AD, n=30; 17 male and 13 female) mice. The APPNL-F/NL-F is a knock-in mouse model of AD, harboring Swedish (APPKM670/671NL) and the Iberian (APPI716F) mutations in APP gene8. APPNL-F mice recapitulate several AD-associated pathologies, including amyloid-β1-42 plaques, synaptic loss, and microgliosis. For MRI/MRS of isoflurane (1.5%) anesthetized mice was performed using a 20mm diameter volume 1H transceiver coil (m2m imaging, USA) in a 30cm horizontal bore 9.4T magnet interfaced with Avance III HD console (Bruker Biospin, Germany). The temperature and respiration of mice were monitored continuously and maintained at 37⁰C and 60-80 beats/minute respectively. The entire imaging protocol consisted of a localizer followed by T1-FLASH (acquisition time = ~4 min), T2-MSME (acquisition time = ~9 min). Anatomical images were used to position our voxel (6x1x2mm3) in the hippocampus and localized water-suppressed 1H MR spectra were acquired (TE=16ms; TR=3000ms). 1H MR spectra were analyzed using the LC Model for the quantification of different metabolites. The terminal methyl group of BCAA (MM09) resonates at ~0.9 ppm which is often overlapped with resonance from the terminal methyl group (Lip 09) of long-chain lipid molecules. The statistical significance of difference across four groups was determined using two-way ANOVA.Results
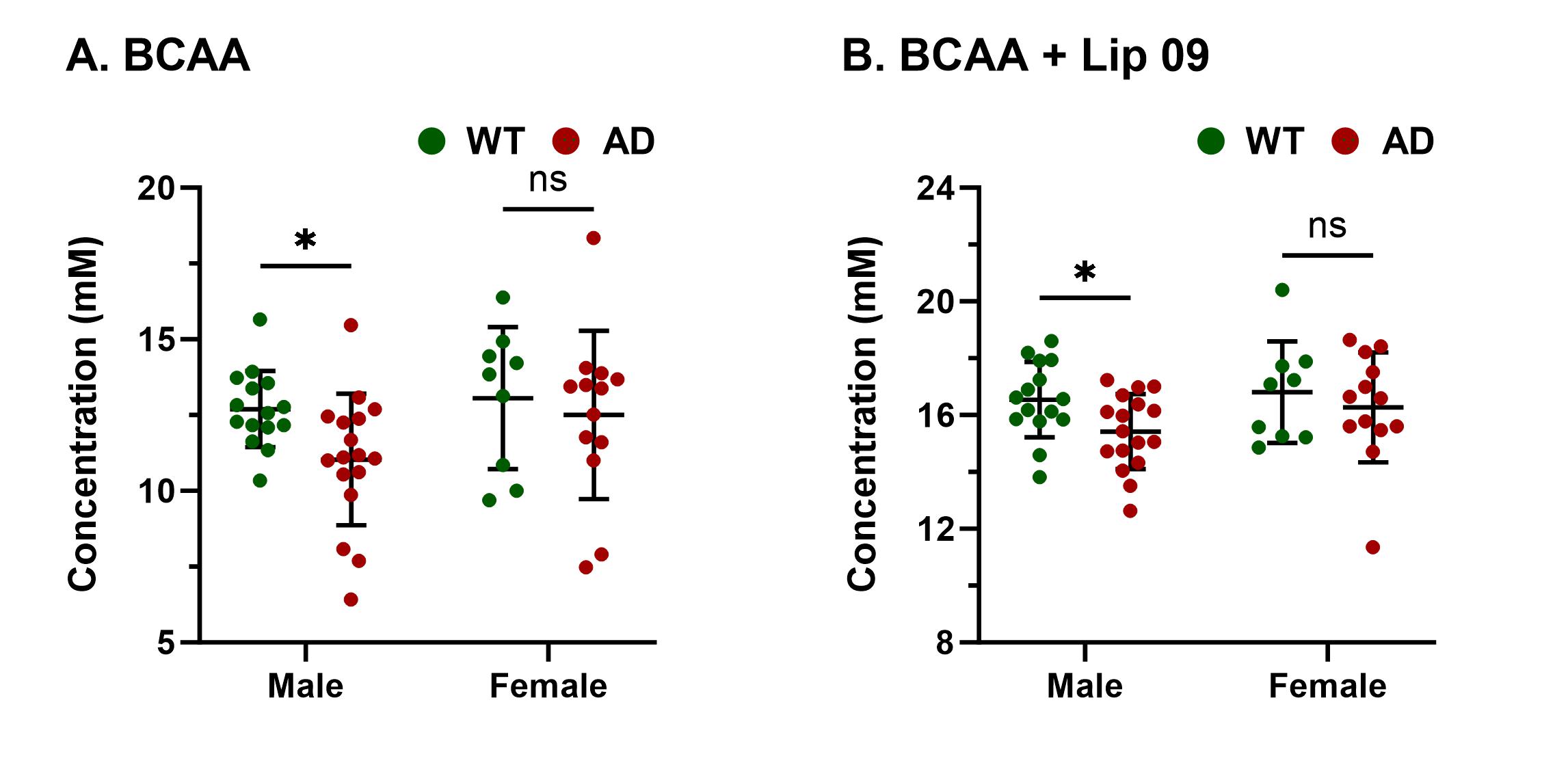
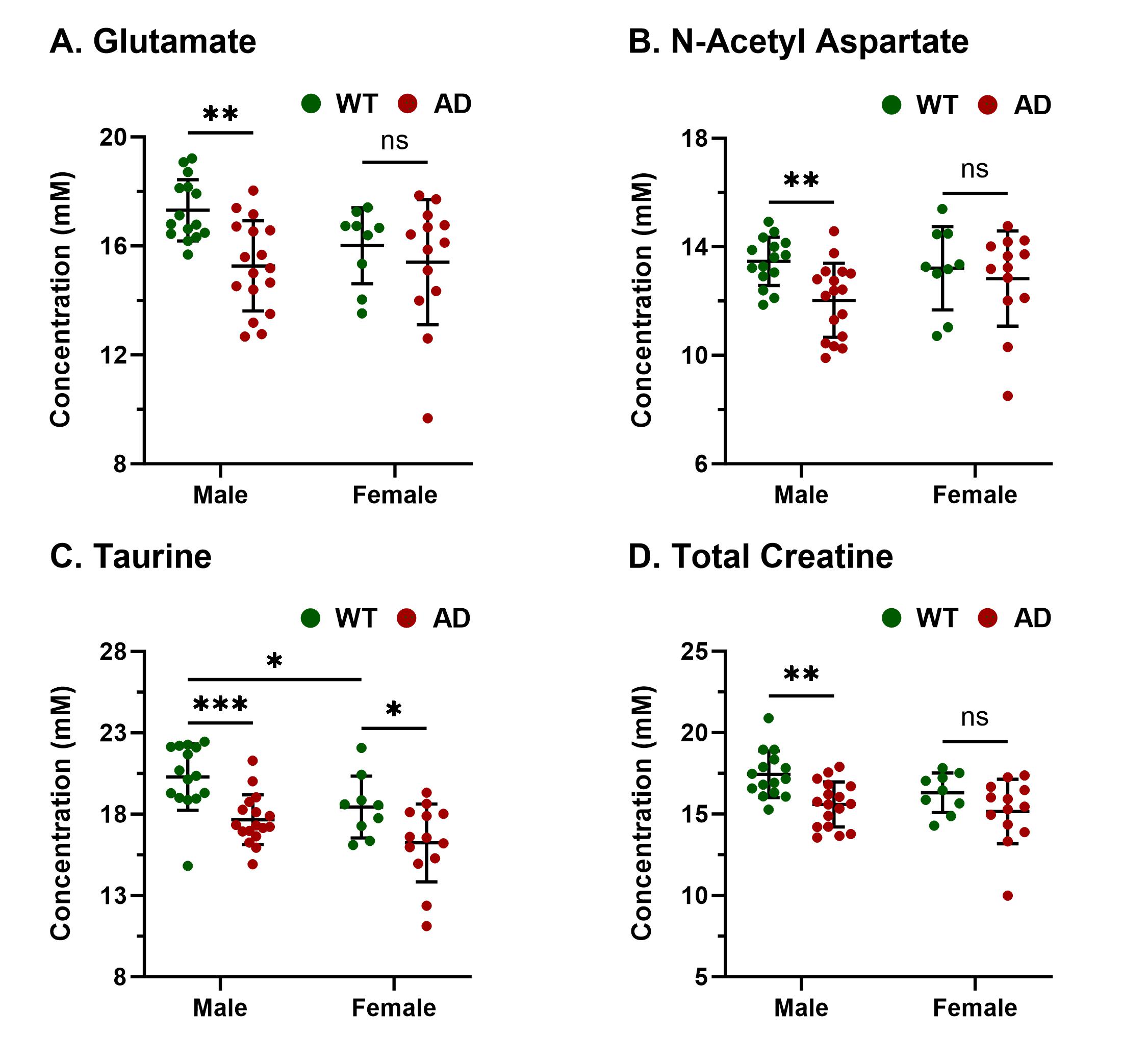
Representative 1H MR spectra from a WT and an AD mouse are shown in Figure 2 where a visible reduction in the level of BCAA, glutamate, and NAA could be seen. Upon quantitative analysis of spectra, we observed a reduction in the levels of BCAA (13%; p=0.03), BCAA+Lip09 (7%; p=0.04) (Fig. 2) glutamate (12%; p=0.001) of male AD mice w.r.t. WT males (Fig. 3). Additionally, these mice also exhibited a loss in the levels of NAA (11%; p= 0.005), taurine (13%; p= 0.0004) and total creatine (tCr 11%; p=0.001) (Fig. 4). However, in the case of female AD mice only taurine (12%; p=0.01) was reduced significantly w.r.t. WT females (Fig. 4). To find any existing association of glutamate levels with BCAA and BCAA+Lip09, we performed correlation analysis and found a significant correlation (p<0.001) with a R2 value of 0.33 and 0.49 respectively (Fig. 5).Discussions
The cerebral cortex and hippocampus are crucial in the regulation of cognitive functions including memory9 and are primarily affected in AD10,11. Our observation of reduced BCAA could be attributed to the replenishment of compromised glutamate and other TCA intermediates by BCAA catabolism. It becomes further evident upon considering the fair correlation between both. Reductions in levels of NAA, taurine, and tCr indicate loss of neurons12, neuroplasticity13, and compromised brain energetics14 due to disrupted Creatine/phospho-Creatine exchange, respectively. Interestingly, female AD mice were almost immune to AD at this stage, which could be mostly attributed to much studied neurotrophic and neuroprotective effects of estrogen15 though it demands a detailed investigation to draw evidence-supported inference. In conclusion, measurement of BCAA using 1H MRS could really help in the development of alternate and specific diagnostic strategies against AD.Acknowledgements
This project was supported by the National Institute of Biomedical Imaging and Bioengineering of the National Institutes of Health through Grant Number P41EB02946 and the National Institute of Aging through Award Number R01AG063869.References
1. Alzheimer's A (2016) 2016 Alzheimer's disease facts and figures. Alzheimers Dement 12:459-509.
2. Polis B, Samson AO. Role of the metabolism of branched-chain amino acids in the development of Alzheimer's disease and other metabolic disorders. Neural Regen Res. 2020 Aug;15(8):1460-1470. doi: 10.4103/1673-5374.274328.
3. Erecinska M and Silver IA (1990) Metabolism and role of glutamate in mammalian brain. Prog Neurobiol 35:245-296.
4. Attwell D and Laughlin SB (2001) An energy budget for signaling in the grey matter of the brain. J Cereb Blood Flow Metab 21:1133-1145.
5. Chen SQ, Cai Q, Shen YY, Wang PJ, Teng GJ, Zhang W and Zang FC (2012) Age-related changes in brain metabolites and cognitive function in APP/PS1 transgenic mice. Behav Brain Res 235:1-6.
6. Patel AB, Tiwari V, Veeraiah P and Saba K (2018) Increased astroglial activity and reduced neuronal function across brain in AβPP-PS1 mouse model of Alzheimer's disease. J Cereb Blood Flow Metab 38:1213-1226.
7. Wang H, Tan L, Wang HF, Liu Y, Yin RH, Wang WY, Chang XL, Jiang T and Yu JT (2015) Magnetic Resonance Spectroscopy in Alzheimer's Disease: Systematic Review and Meta-Analysis. J Alzheimers Dis 46:1049-1070.
8. Saito, T., Matsuba, Y., Mihira, N., Takano, J., Nilsson, P., Itohara, S., Iwata, N., & Saido, T. C. (2014). Single App knock-in mouse models of Alzheimer's disease. Nature neuroscience, 17(5), 661–663.
9. Ito, I., & Sugiyama, H. (1991). Roles of glutamate receptors in long-term potentiation at hippocampal mossy fiber synapses. Neuroreport, 2(6), 333–336
10. Jack CR, Jr., Petersen RC, Xu Y, O'Brien PC, Smith GE, Ivnik RJ, Boeve BF, Tangalos EG and Kokmen E (2000) Rates of hippocampal atrophy correlate with change in clinical status in aging and AD. Neurology 55:484-489.
11. Pearson RC, Esiri MM, Hiorns RW, Wilcock GK and Powell TP (1985) Anatomical correlates of the distribution of the pathological changes in the neocortex in Alzheimer disease. Proc Natl Acad Sci USA 82:4531-4534.
12. Rigotti, D. J., Inglese, M., & Gonen, O. (2007). Whole-brain N-acetylaspartate as a surrogate marker of neuronal damage in diffuse neurologic disorders. AJNR. American journal of neuroradiology, 28(10), 1843–1849. https://doi.org/10.3174/ajnr.A0774
13. Rafiee Z, García-Serrano AM, Duarte JMN (2022). Taurine Supplementation as a Neuroprotective Strategy upon Brain Dysfunction in Metabolic Syndrome and Diabetes. Nutrients. 2022 Mar 18;14(6):1292.
14. Forbes SC, Cordingley DM, Cornish SM, Gualano B, Roschel H, Ostojic SM, Rawson ES, Roy BD, Prokopidis K, Giannos P, Candow DG (2022). Effects of Creatine Supplementation on Brain Function and Health. Nutrients. 2022 Feb 22;14(5):921.
15. Brann DW, Dhandapani K, Wakade C, Mahesh VB, Khan MM (2007). Neurotrophic and neuroprotective actions of estrogen: basic mechanisms and clinical implications. Steroids. 2007 May;72(5):381-405.
Figures