0739
IL-6 knockout reduces doxorubicin-induced neurotoxicity1Translational Medicine, The Hospital for Sick Children, Toronto, ON, Canada, 2Department of Medical Biophysics, University of Toronto, Toronto, ON, Canada, 3Departments of Paediatrics, The Hospital for Sick Children, Toronto, ON, Canada, 4Psychology Department, The Hospital for Sick Children, Toronto, ON, Canada, 5Department of Psychiatry, The Hospital for Sick Children, Toronto, ON, Canada, 6Division of Clinical Pharmacology & Toxicology, The Hospital for Sick Children, Toronto, ON, Canada
Synopsis
Keywords: Neuroinflammation, Cancer, chemotherapy, treatment, late effects, pediatric, development
Motivation: Doxorubicin (DXR) is a widely used chemotherapy agent associated with inflammation and neurocognitive impairment in cancer survivors. Given that DXR has limited access to the brain, indirect mechanisms, such as the generation of systemic pro-inflammatory cytokines, are proposed to induce neurotoxicity and neuroinflammation.
Goal(s): This study aims to probe this hypothesized pro-inflammatory pathway of DXR-induced neurotoxicity.
Approach: We identified an elevation of the pro-inflammatory cytokine IL-6 in DXR-treated mice. Consequently, we utilized MRI to assess neuroanatomical changes after DXR treatment in wildtype and Il-6 knockout mice.
Results: Our findings revealed that Il-6 knockout partially mitigated the neurotoxic effects induced by DXR.
Impact: DXR leads to cognitive impairment that diminishes quality of life for cancer survivors. We demonstrated the involvement of IL-6 in the neurotoxic mechanism of DXR, suggesting a strategy for targeting IL-6 to limit neurotoxicity of cancer treatments.
Introduction
Doxorubicin (DXR) is one of the most widely used chemotherapeutic agents for cancer treatment. While DXR is an effective anti-cancer drug, it can be responsible for organ toxicity and late side effects1,2. A study using a mouse model for pediatric leukemia treatment demonstrated that DXR – similar to a couple of other chemotherapeutics agents – induces widespread volume deficits in the brain3. However, DXR has limited penetrance through the blood-brain barrier, so its neurotoxicity mechanism is likely indirect. DXR-induced systemic inflammation is one such indirect mechanism, in which elevated pro-inflammatory cytokines levels generated systemically are hypothesized to pass into the brain and mediate neuroinflammatory response4. To investigate this mechanism, we utilized a multiplex cytokine panel to identify cytokine elevations that may contribute to systemic inflammation after DXR treatment, revealing significant increases in IL-6. We used in vivo structural MRI to show that the knockout of Il-6 reduces long-term DXR-induced neurotoxicity in several affected brain regions by assessing brain volumes.Methods
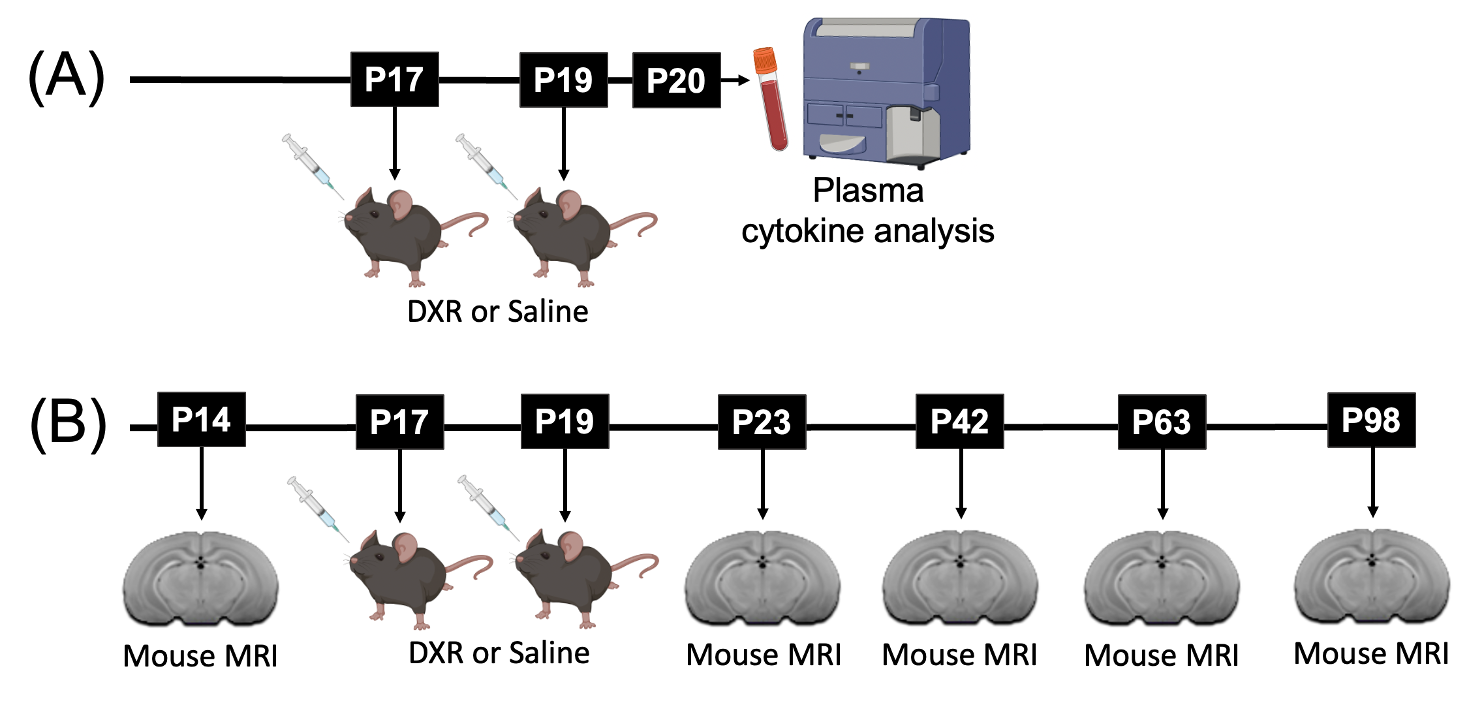
To screen for cytokines in the blood, C57BL/6J wildtype mice were treated with DXR or saline treatment at postnatal day (P)17 and P194. The mice were sacrificed at P20 to extract plasma which was then analyzed using the Luminex 25-plex assay (Figure 1A). For longitudinal MRI, F1 hybrids of CD1 and C57BL/6J mice with a homozygous knockout of Il-6 or wildtype Il-6 received DXR or saline treatment at P17 and P19 (n=12 per group). Mice were scanned at P14, P23, P42, P63, and P98 (Figure 1B). In vivo images with enhanced manganese contrast were obtained using a 3D T1-weighted gradient echo sequence on a multi-channel 7T MRI scanner5. The acquisition parameters included a 75μm isotropic resolution, TR of 26ms, TE of 8.25ms, a flip angle of 26°, a field-of-view measuring 25×22×22mm, and a matrix size of 334×294×294. Subsequently, automated registration techniques were employed to segment the images into 183 distinct structures.6,7 A linear mixed-effects model was used for structure-specific analysis.Results
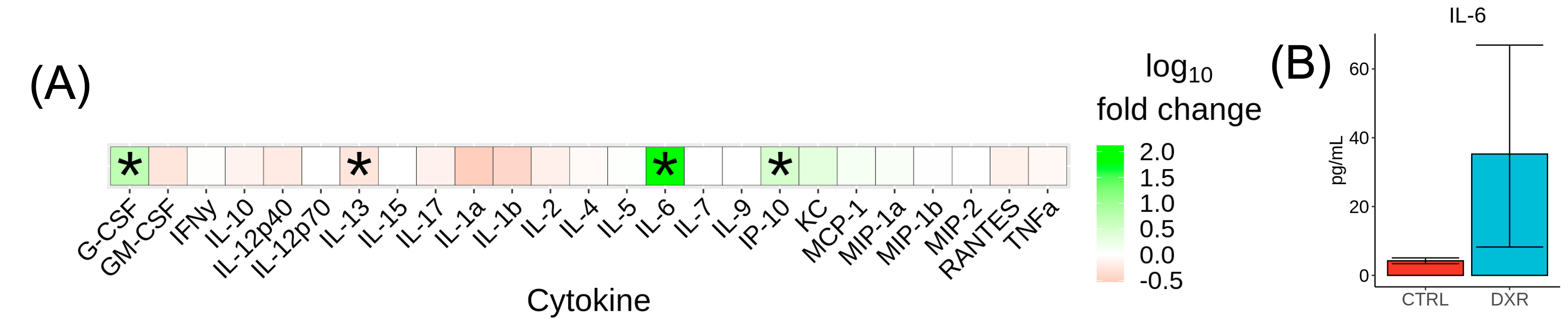
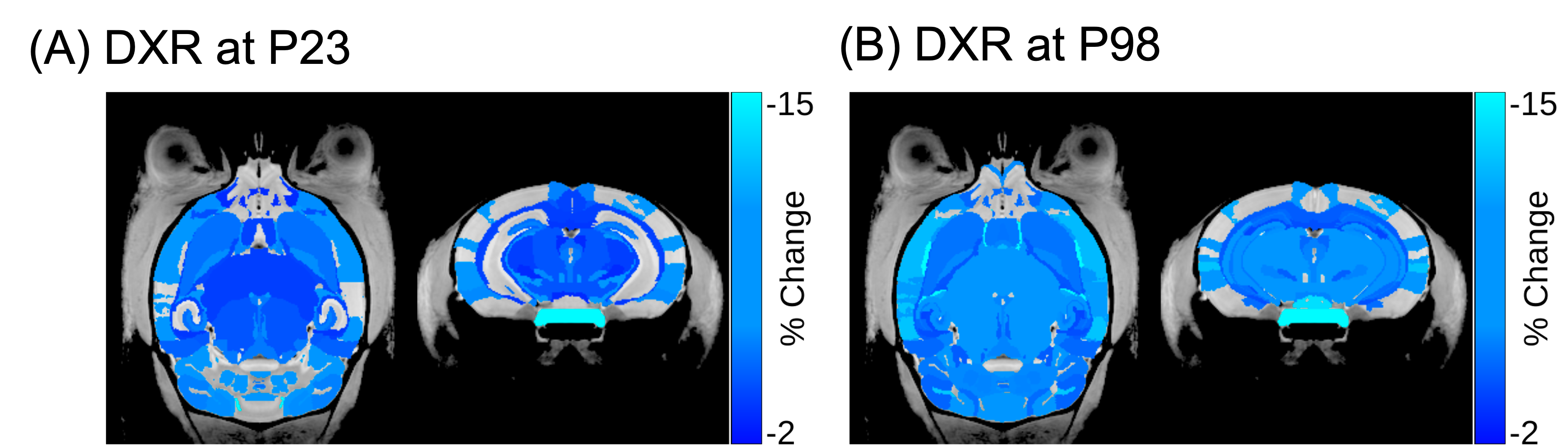
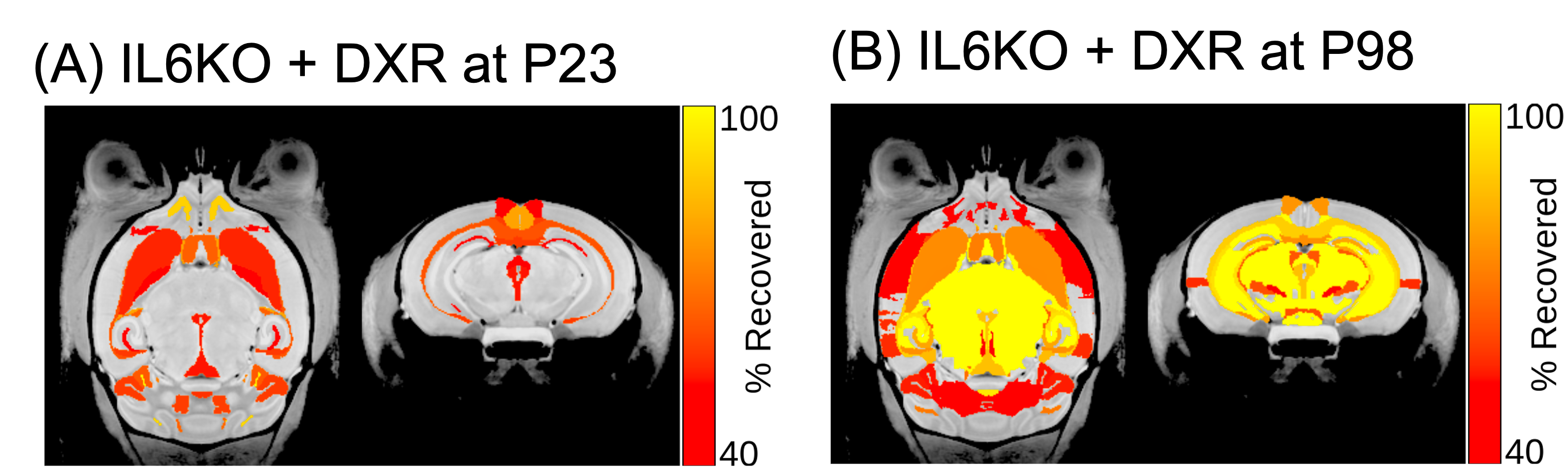
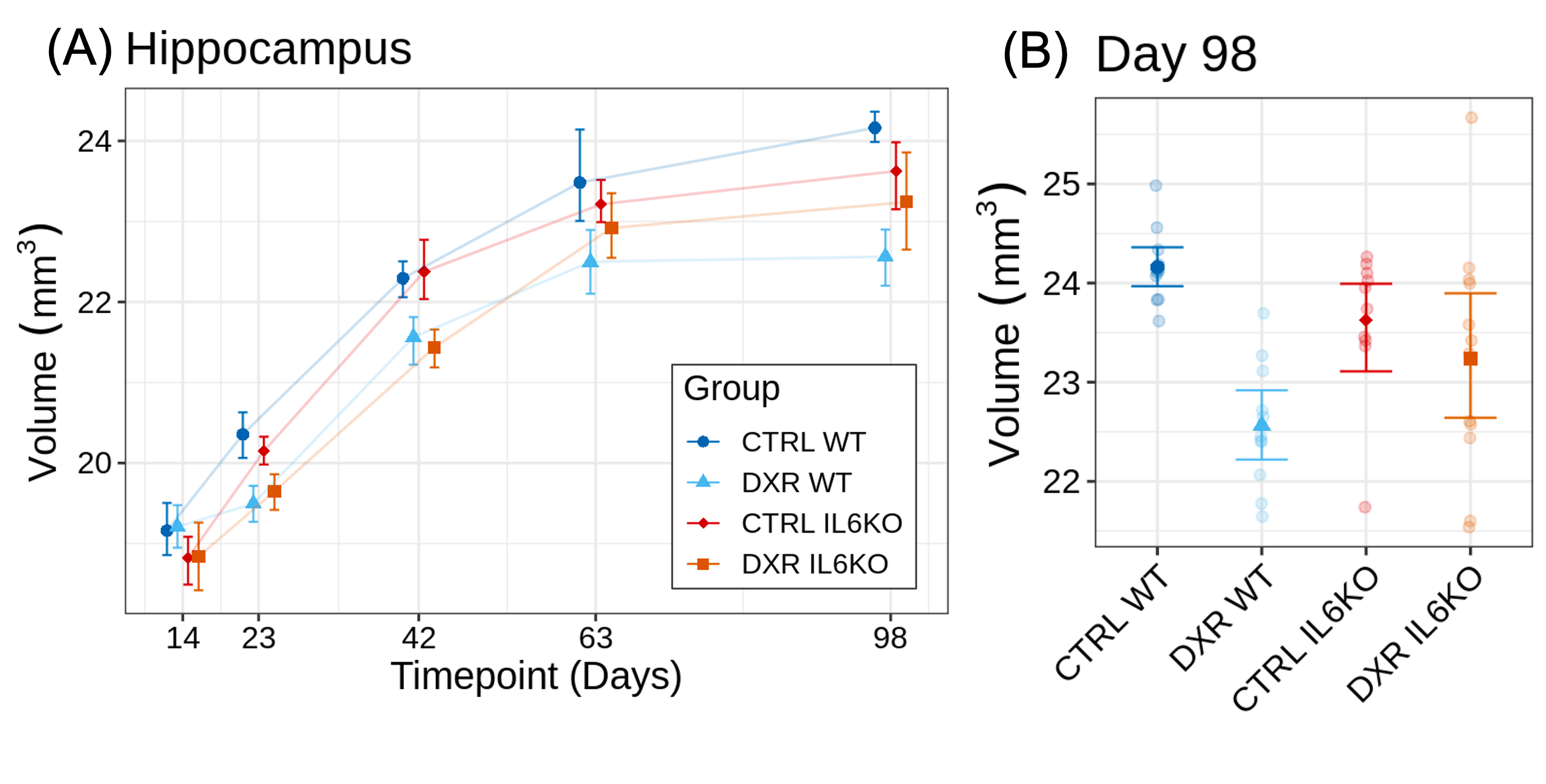
The cytokine panel revealed statistically significant alterations in G-CSF, IL-13, IP-10, and IL-6 (Figure 2A). The cytokine IL-6 was found to be most significantly elevated in the DXR group compared to controls (131-fold increase, p=0.026) (Figure 2B). Figure 3 shows the DXR-induced widespread volume loss in the brain at P23 (99 out of 183 structures) and P98 (105 out of 183) that were statistically significant (FDR<10%). Knockout of Il-6 resulted in improved volume outcomes after DXR treatment (Figure 4). Twice as many structures exhibited at least a 40% improvement in outcome at P98 (62 out of 183) as at P23 (31 out of 183). Some of the recovered structures include the hippocampus (Figure 5), midbrain, pons, and corpus callosum.Discussion
DXR treatment is known to cause systemic inflammation and neurocognitive deficits8,9. The identification of systemic IL-6 elevation in DXR-treated mice is consistent with the known role of IL-6 in mediating systemic inflammation and the febrile response10. While DXR does not efficiently enter the brain, circulating IL-6 can pass through the blood-brain barrier and induce neuroinflammation, contributing to neurotoxicity8 . Our observations support the involvement of IL-6 in mediating toxicity as knockout of Il-6 reduced DXR-induced volume loss. We also observed enhanced recovery at later timepoints, suggesting that IL-6 may influence brain repair mechanisms during the later phases post-treatment. To further understand the role of IL-6, additional experiments will be required to determine whether systemic or central nervous system production of IL-6 is most important, and the role of IL-6 in the acute versus chronic phases post-treatment.Conclusion
DXR treatment has been linked to neurocognitive deficits in cancer survivors. To investigate possible mechanisms responsible for DXR-induced neurotoxicity, we evaluated cytokine elevations in DXR-treated mice and identified IL-6 as a candidate. We used longitudinal MRI to demonstrate a reduction in DXR-induced neurotoxicity in Il-6 knockout mice. These results provide important insight on possibly targeting pro-inflammatory pathways that involve IL-6 to mitigate neurocognitive deficits in cancer survivors treated with DXR.Acknowledgements
This work is supported in part by the Canadian Institute of Health Research, the Ontario Institute for Cancer Research, and a Restracomp award administered by the SickKids Research Training Centre.References
1. Carvalho, C. et al. Doxorubicin: the good, the bad and the ugly effect. Curr Med Chem 16, 3267–3285 (2009).
2. Wang, L. et al. Doxorubicin-Induced Systemic Inflammation Is Driven by Upregulation of Toll-Like Receptor TLR4 and Endotoxin Leakage. Cancer Res 76, 6631–6642 (2016).
3. Spencer Noakes, T. L. et al. Brain Development and Heart Function after Systemic Single-Agent Chemotherapy in a Mouse Model of Childhood Leukemia Treatment. Clin Cancer Res 24, 6040–6052 (2018).
4. Du, J. et al. Doxorubicin-Induced Cognitive Impairment: The Mechanistic Insights. Front Oncol 11, 673340 (2021).
5. Arbabi, A. et al. Multiple-mouse magnetic resonance imaging with cryogenic radiofrequency probes for evaluation of brain development. NeuroImage 252, 119008 (2022).
6. Friedel, M., van Eede, M. C., Pipitone, J., Chakravarty, M. M. & Lerch, J. P. Pydpiper: a flexible toolkit for constructing novel registration pipelines. Front. Neuroinform. 8, (2014).
7. Chakravarty, M. M. et al. Performing label-fusion-based segmentation using multiple automatically generated templates: MAGeT Brain: Label Fusion Segmentation Using Automatically Generated Templates. Hum. Brain Mapp. 34, 2635–2654 (2013).
8. Wang, X.-M. et al. Chemobrain: A critical review and causal hypothesis of link between cytokines and epigenetic reprogramming associated with chemotherapy. Cytokine 72, 86–96 (2015).
9. Eide, S. & Feng, Z.-P. Doxorubicin chemotherapy-induced ‘chemo-brain’: Meta-analysis. Eur J Pharmacol 881, 173078 (2020).
10. Tanaka, T., Narazaki, M. & Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb Perspect Biol 6, a016295 (2014).
Figures