0702
Metabolic and imaging phenotypes associated with RB1 loss in castrate resistant prostate cancer13. Laboratory of Genitourinary Cancer Pathogenesis, NCI/NIH, Bethesda, MD, United States, 2Laboratory of Genitourinary Cancer Pathogenesis, NCI/NIH, Bethesda, MD, United States, 3Molecular Imaging Branch, NCI/NIH, Bethesda, MD, United States, 4Urologic Oncology Branch, NCI/NIH, Bethesda, MD, United States, 56. Clinical Research Directorate, Frederick National Laboratory for Cancer Research, NCI/NIH, Frederick, MD, United States, 6Radiation Biology Branch, NCI/NIH, Bethesda, MD, United States
Synopsis
Keywords: Biology, Models, Methods, Cancer, prostate, metabolomics
Motivation: The progression of prostate cancer is marked by both RB1 and T53 inactivation and higher 18FDG-PET uptake, but it's unclear whether RB1 or TP53 inactivation drives increased glucose import.
Goal(s): Can metabolic changes be used as a biomarker for RB1 and TP53 loss?
Approach: Metabolomic analysis by NMR and IC-MS for a comprehensive measure of metabolic changes ex vivo and and hyperpolarized MRI to measure the Warburg effect in vivo.
Results: 18FDG uptake was unaffected by loss of either RB1 or TP53. RB1 and TP53 did induce a series of other metabolic changes which could be detected in vivo by hyperpolarized MRI
Impact: Neuroendrocrine prostate cancer is a life-threatening progression of prostate cancer that is characterized by mutations in two key genes. Hyperpolarized MRI may enhance early diagnosis of NEPC without biopsy.
Introduction
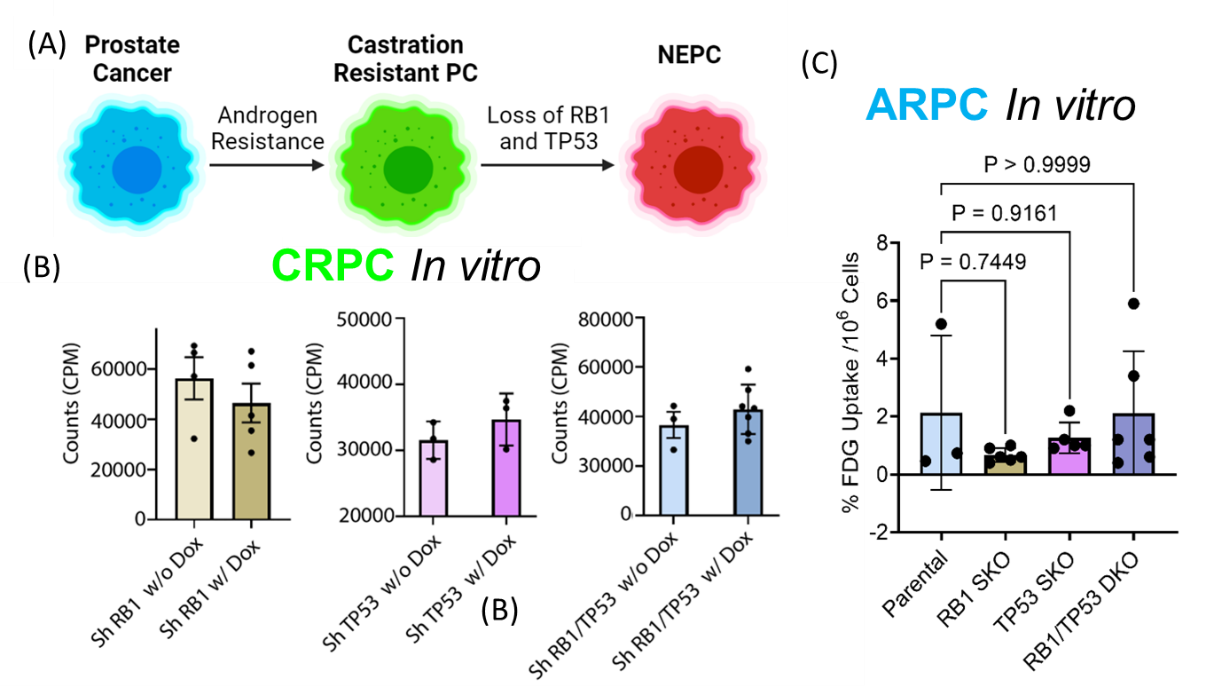
Prostate cancer develops in a series of stages (Fig. 1A). While initially dependent on androgen receptor signaling,(1) about 15-20% patients eventually lose dependence on androgen receptor signaling and develop potentially lethal castrate resistant prostate cancer (CRPC).(1,2) Loss of RB1 in CRPC tumors is correlated with rapid progression and poor patient survival,(3) and in combination with TP53 loss,(4,5) predisposes to the development of lethal transitional neuroendocrine prostate cancer (NEPC) (Fig. 1A). Although the progression of CRPC to NEPC is clinically associated with higher 18FDG -PET SUVmax values,(6) it is unknown whether inactivation of RB1 and/or TP53 drives this increased glucose import.Methods
Knockdown models of RB1/TP53 loss in CRPC were created by doxycycline induction of short hairpin RNA in an established organoid model of CRPC (LuCaP 167) in which RB1 and TP53 signaling are intact. Single and double knockouts of RB1/TP53 in androgen receptor signaling positive prostate cancer (ARPC) were created by CRISPR in an ARPC cell line (LnCaP). Hyperpolarized 13C MRI experiments were performed on a 3T MRI Scanner (MR Solutions Inc.) using a 17-mm diameter home-built 1H/13C coil. with a CSI imaging sequence with a 32 × 32 mm field of view in an 8-mm axial slice.RB1/TP53 knockdown does not affect 18FDG uptake in models of CRPC or ARPC
No significant difference in FDG activity was detected after RB1, TP53, or RB1/TP53 knockdown in either LuCaP 167 mouse xenograft models of CRPC in vivo (Fig. 1C) or in LnCaP cellular models of ARPC (Fig. 1D) in vitro. Hyperpolarized 13C MRI experiments were performed on a 3T MRI Scanner (MR Solutions Inc.) using a 17-mm diameter home-built 1H/13C coil. with a CSI imaging sequence with a 32 × 32 mm field of view in an 8-mm axial slice.Ex vivo metabolomics shows combined knockdown of RB1/TP53 leads to metabolic changes, including a diversion of glucose into glycogenesis
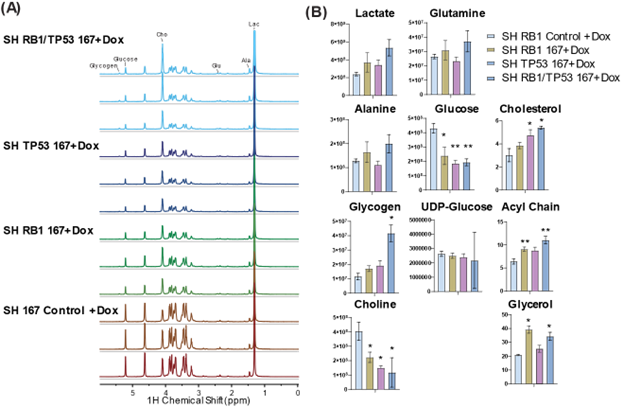
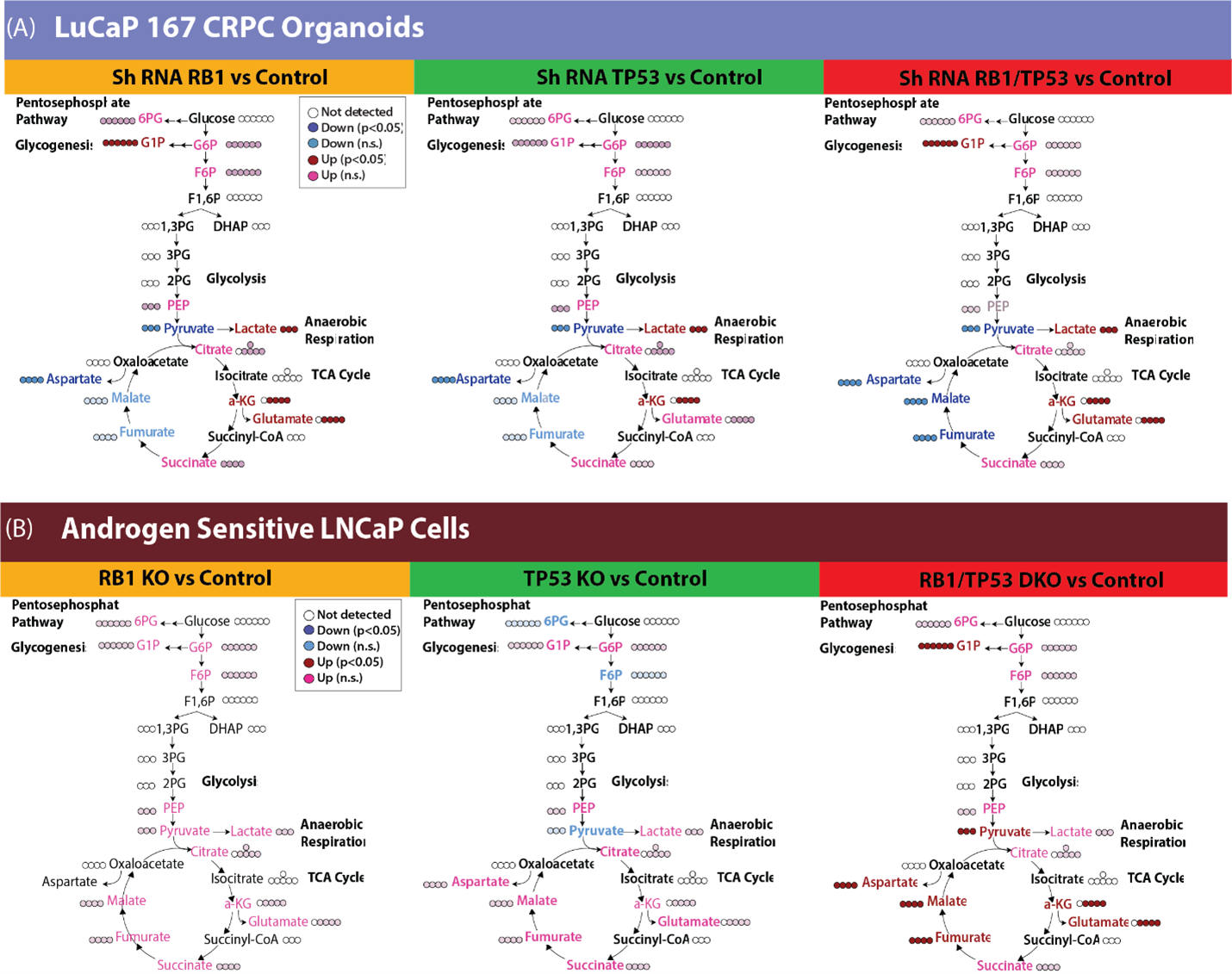
FDG-PET is limited to measuring glucose uptake and phosphorylation. To gain a more comprehensive understanding of the metabolic transformation after RB1 and/or TP53 depletion, we quantified the downstream metabolites of 13C glucose in the LuCaP 167 CRPC organoid model by NMR (Fig 2). 13C glycogen was highly enriched after dual RB1/TP53 depletion while steady state concentrations of 13C glucose decreased, suggesting a diversion of glucose to form glycogen. Consistent with Seahorse and LDH activity assays, 13C labeled lactate concentrations were elevated after dual RB1/TP53 depletion with more modest increases occurring following single gene modifications. In the non-polar fraction, both cholesterol synthesis and 13C incorporation into lipid acyl chains and glycerol headgroups were significantly elevated after depletion of either RB1 or TP53. The androgen sensitive LnCaP model was less affected relative to LuCaP167 by RB1/TP53 loss when glucose metabolism was measured by 13C NMR. We therefore repeated the 13C glucose tracer experiment using targeted IC-MS to detect changes in intermediates in the glycolytic and TCA pathways whose low abundance is problematic for NMR (Fig. 3). In the LuCaP 167 organoid CRPC models, RB1 and TP53 depletions were clearly differentiated from control samples by IC-MS primarily by a decrease in pyruvate and an increase in lactate concentrations. In LnCAP ARPC models, the primary difference was observed in the combined RB1/TP53 null samples. Dual RB1/TP53 knockout increased activity throughout the TCA cycle, which was not seen to a significant extent with individual knockouts. To summarize, RB1 loss in different ARPC models led to consistent changes specifically, 1) increased lactate, reflecting higher lactate dehydrogenase activity, 2) increased glucose-1-phosphate and glycogen levels, suggesting redirection of glucose metabolism to gluconeogenesis, and 3) modulation of the TCA cycle, specifically a consistent increase in αKG across models in addition to other model-specific changes.Increase in LDH flux after RB1 knockdown can be detected in vivo by 13C-HPMRS
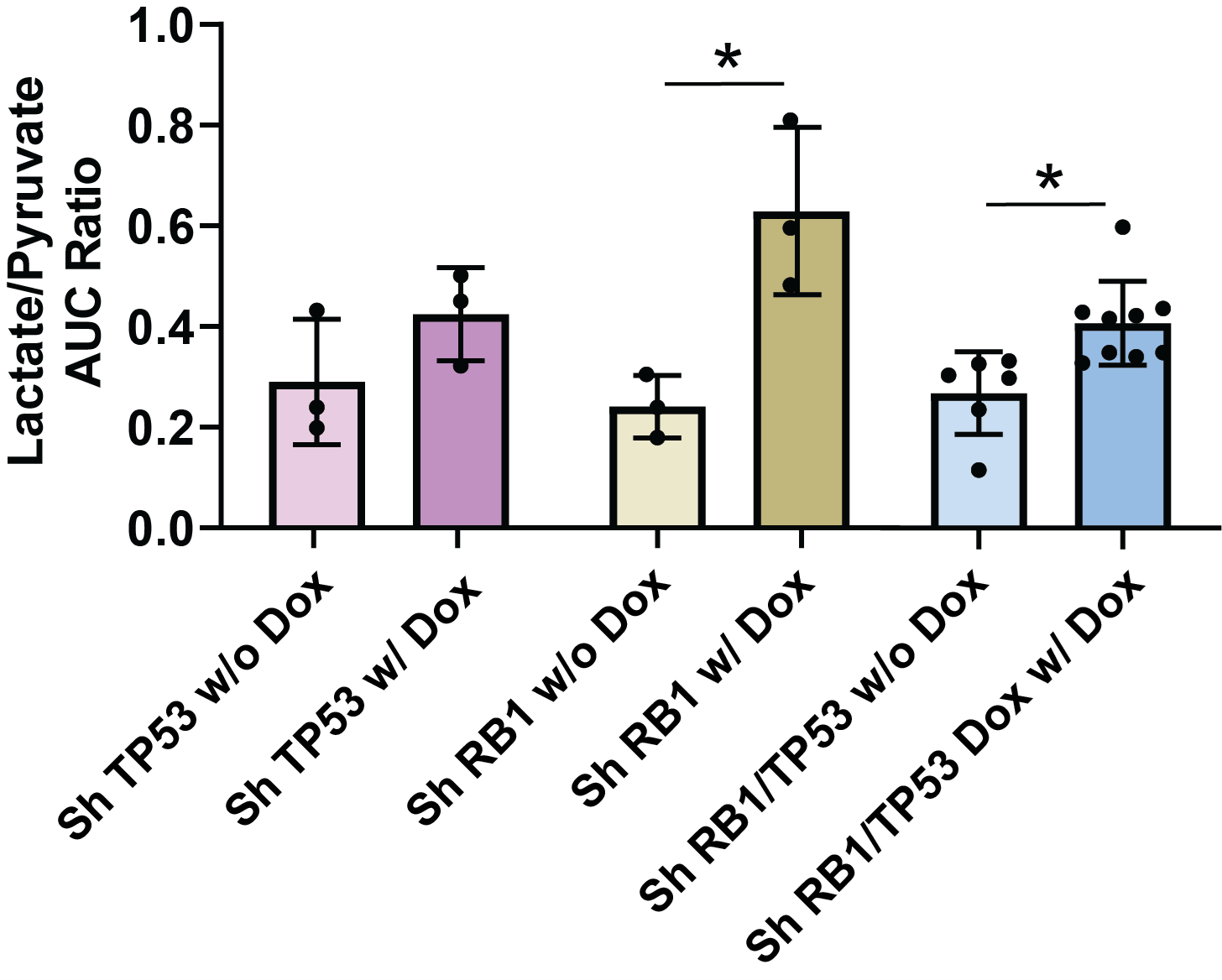
NMR and ICMS identified several metabolites that were upregulated after RB1/TP53 knockdown. To determine whether such a phenotype could be detected in vivo, we assayed LuCaP167 organoid-derived PDX tumors in vivo (Fig. 4) using hyperpolarized MRI to detect the de novo generation of new metabolites from pyruvate. In LuCaP 167 CRPC tumors, the rate of pyruvate to lactate conversion significantly increased with depletion of RB1 with or without partial depletion of TP53.Conclusion
Functional loss of RB1 is the most significant predictive mutation for poor survival in CRPC (1). While higher 18FDG -PET SUVmax values are also associated with tumor aggressiveness and poor survival in CRPC, in our models of both CRPC and ARPC, RB1 loss is not associated with increased 18FDG uptake. Instead, we found that both RB1 and TP53 are associated with extensive metabolic reprogramming that can be detected ex vivo by NMR and in vivo by hyperpolarized MRI.Acknowledgements
This project has been funded in whole or in part with federal funds from the National Cancer Institute, National Institutes of Health, under Contract No. 75N91019D00024. The content of this publication does not necessarily reflect the views or policies of the Department of Health and Human Services, nor does mention of trade names, commercial products, or organizations imply endorsement by the U.S. Government.References
1. Freedland SJ, Humphreys EB, Mangold LA, et al. Risk of Prostate Cancer–Specific Mortality Following Biochemical Recurrence After Radical Prostatectomy. JAMA. 2005;294:433-439.
2. Watson PA, Arora VK, Sawyers CL. Emerging mechanisms of resistance to androgen receptor inhibitors in prostate cancer. Nat Rev Cancer. 2015;15:701-711.
3. Abida W, Cyrta J, Heller G, et al. Genomic correlates of clinical outcome in advanced prostate cancer. Proc Natl Acad Sci U S A. 2019;116:11428-11436.
4. Beltran H, Prandi D, Mosquera JM, et al. Divergent clonal evolution of castration-resistant neuroendocrine prostate cancer. Nat Med. 2016;22:298-305.
5. Sheng Yu Ku SR, Yanqing Wang, Ping Mu, Mukund Seshadri,, Zachary W. Goodrich MMG, David P. Labbé,, Eduardo Cortes Gomez JW, Henry W. Long, Bo Xu, Myles Brown,, Massimo Loda CLS, Leigh Ellis, David W. Goodrich. Rb1 and Trp53 cooperate to suppress prostate cancer lineage plasticity, metastasis, and antiandrogen resistance. Science. 2017;355:78-83.
6. Spratt DE, Gavane S, Tarlinton L, et al. Utility of FDG-PET in clinical neuroendocrine prostate cancer. Prostate. 2014;74:1153-1159.
Figures