0694
Using a standard MRI atlas with non-equitable sex representation introduces spurious sex effects1Instituto de Neurociencias, CSIC-UMH, Alicante, Spain
Synopsis
Keywords: Preclinical Image Analysis, Data Analysis, template, Allen brain
Motivation: Overcoming sex bias in preclinical research requires unbiased tools. Equitable sex representation in the creation of the atlas is seldom achieved, but failing to do so might introduce bias in the analysis.
Goal(s): Our goal was to create a mouse MRI template that includes animals of both sexes, incorporating both structural and diffusion contrasts, and then compare it with a single-sex template.
Approach: We scanned a large cohort of mice, including both sexes, to generate a template and registered the Allen annotation to it.
Results: Using single-sex templates generates spurious sex differences that are not present when using resources with equitable sex representation.
Impact: The MouseX DW-Allen Atlas is the first mouse MRI template that adopts the Allen annotation and is built with both sexes. It fills an important gap in preclinical imaging research, promotes gender equality, and fosters collaborative and multi-approach science.
INTRODUCTION
Sexual dimorphism has a prominent role in brain physiological and pathological trajectories1,2,3. While efforts have been made to overcome sex-bias in clinical trials, progress has been limited in preclinical research. Only few standard atlases for mouse microstructural MR imaging are available, and none of them include both sexes. Therefore, our aim was to i) create a new resource facilitating mouse microstructural MRI and its combination with other content already available in the Allen brain database, and ii) test whether the use of an atlas generated with a single sex introduces bias in the data analysis.METHODS
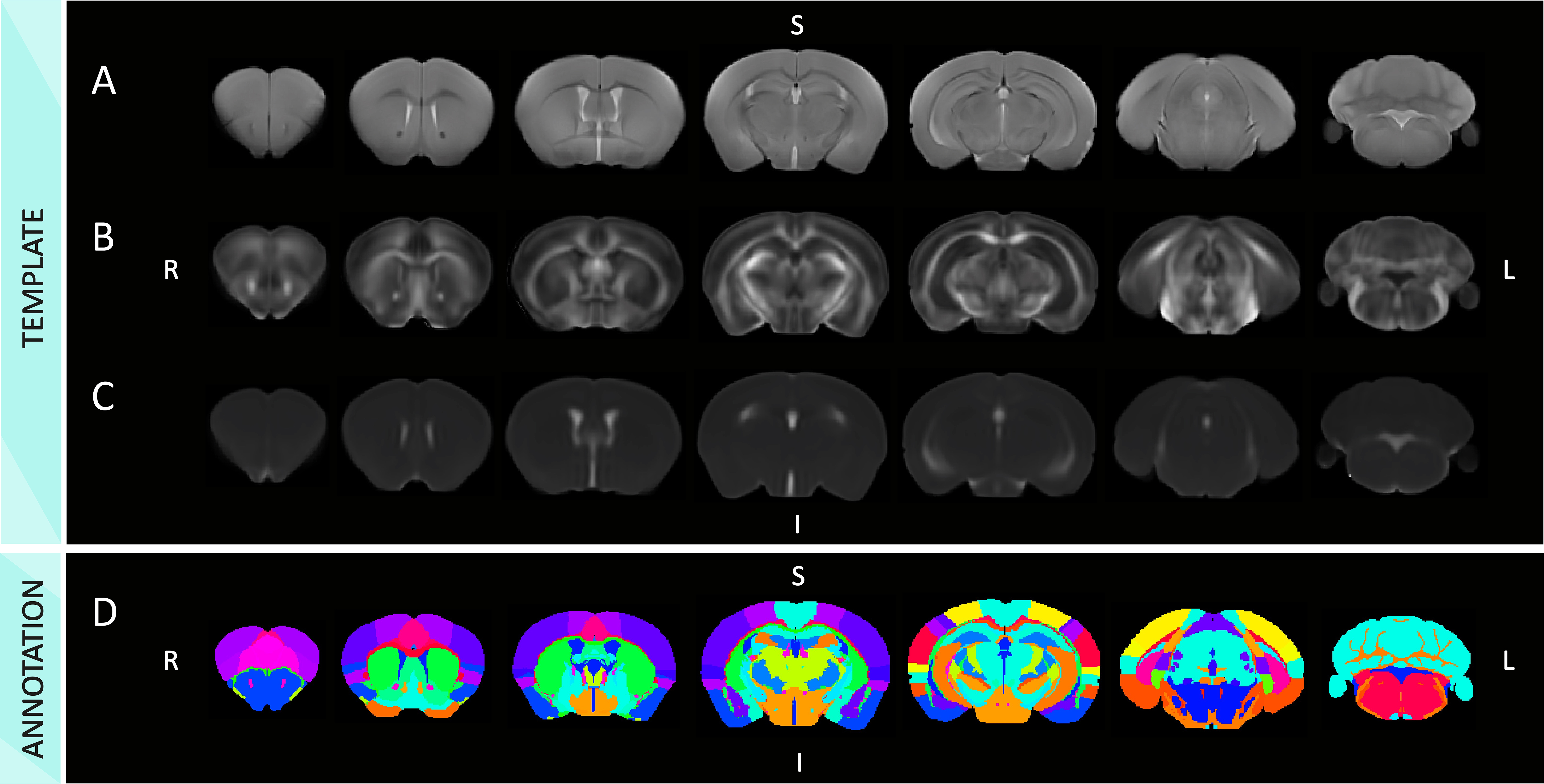
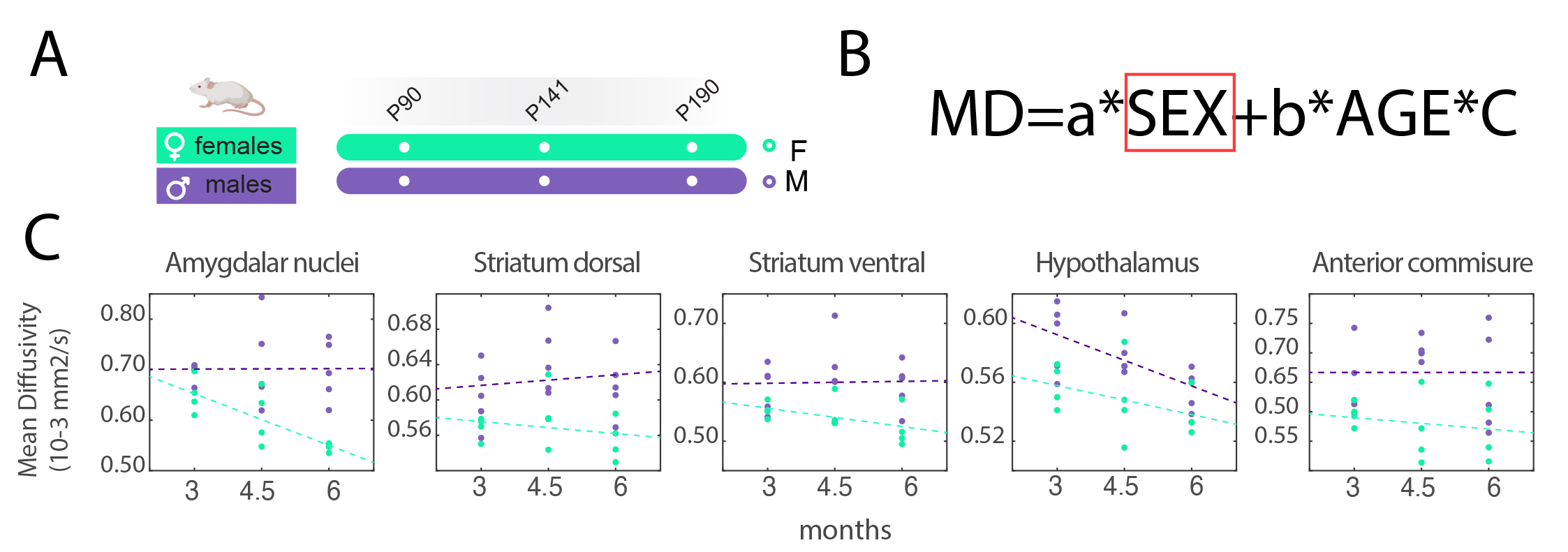
MRI experiments were conducted on a Bruker 7T animal scanner. On the first cohort of mice (n=60), T2-Weighted (T2W) data was acquired with a RARE sequence, matrix size=258x214, in-plane voxel size=70µm2, forty slices of thickness=400µm, TR=3394ms, TE=33.5ms. In forty of these animals (20 females), a diffusion-weighted (dw) sequence was acquired with 20 gradient orientations, b=1000 s/mm2, diffusion time 15ms, four b0s, TR=8000ms, TE=26ms. On the second cohort of animals (n= 9, 4 females) T2W data was acquired longitudinally with a RARE sequence, matrix size=108x90, in-plane voxel size=160µm2, sixteen slices of thickness=800µm, TR=3000ms, TE=7.7ms. Dw-MRI data were acquired with 20 gradient orientations, diffusion time 15ms, b=1000 s/mm2, four b0s, TR=8000ms, and TE=25ms.MRI raw data was processed using ANTs5 for image registrations and FSL BET6 for skull removal. Dw-MRI data was corrected for EPI distortions, motion distortions, and free water7; data was denoised using patch2self8 and fractional anisotropy (FA) and mean diffusivity (MD) maps were extracted using ExploreDTI9. Both T2W and dw-MRI maps were separately processed for the template generation using ANTs tool buildtemplateparallel.sh. Two-photon tomography-based AMBA-CCFv3 annotation and template10 served as a starting point, and was non-linearly registered to the T2W template using affine and SyN transformation. Affine and inverse warp transforms were applied to the AMBA-CCFv3 annotation to bring it in the MRI space, revised and corrected voxel-wise, following Allen annotation. Parcellation was reduced to 63 regions-of-interest (ROI) to match MRI resolution more accurately. The atlas was named MouseX DW-ALLEN and was uploaded online.To demonstrate the potential of sharing a common annotation across neuroimaging techniques, we tested for correlations between ROI-wise cell type counting form the 3D Cell-Atlas for the Mouse Brain delineated by Erö et al11and MD. P-values values were corrected for multiple comparisons12. Voxel-wise comparison between deformation fields in males and females to match the MouseX DW-ALLEN template, and its male-only version, was evaluated by comparing the Jacobian determinant across sexes13. In the second cohort scanned at three time points, the two templates (both sexes and male-only) were used to extract the average MD values per region, and a general linear model was tested for each ROI, with age and sex as factors. P-values were corrected for multiple comparisons using false discovery rate.
RESULTS
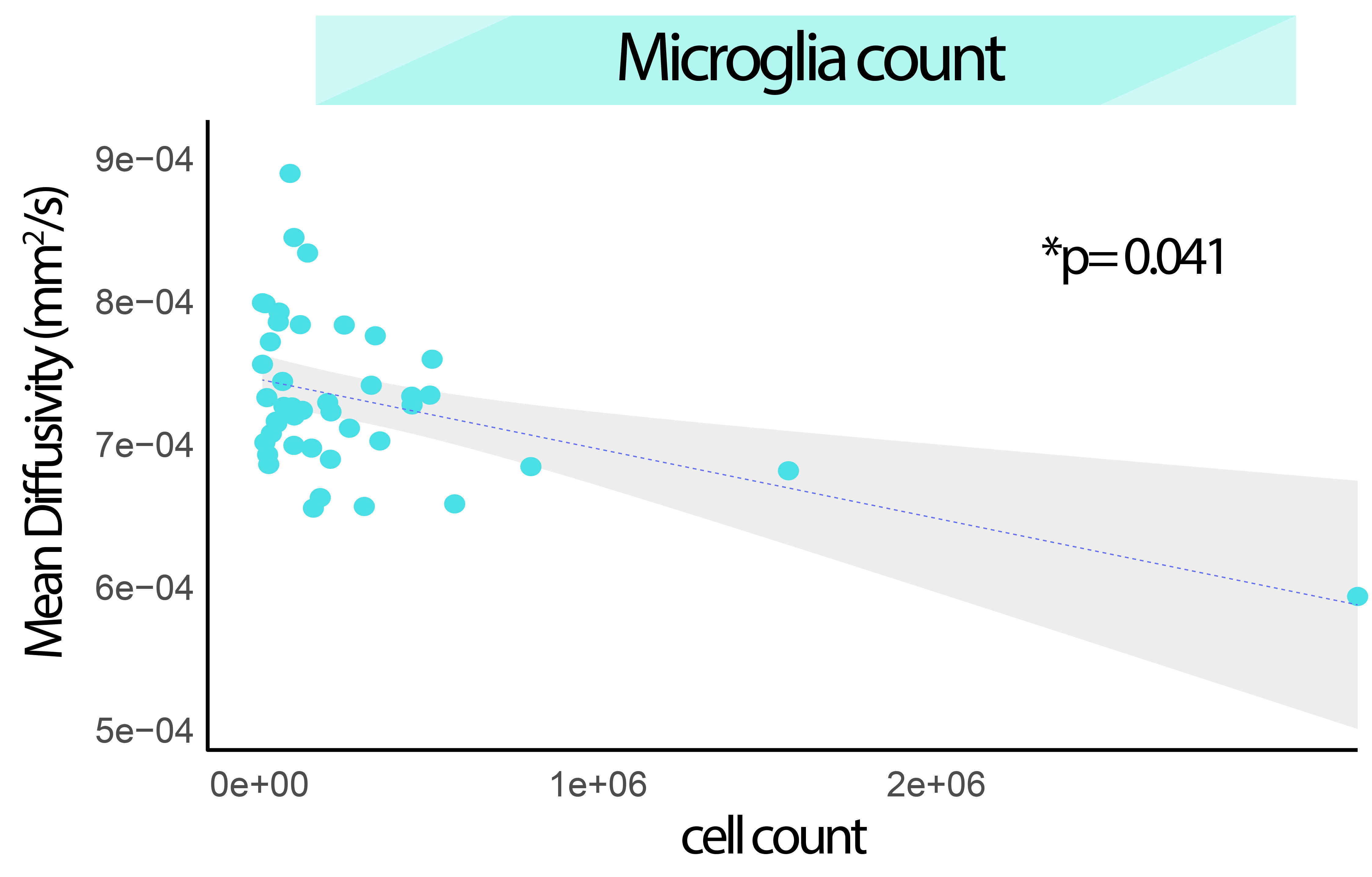
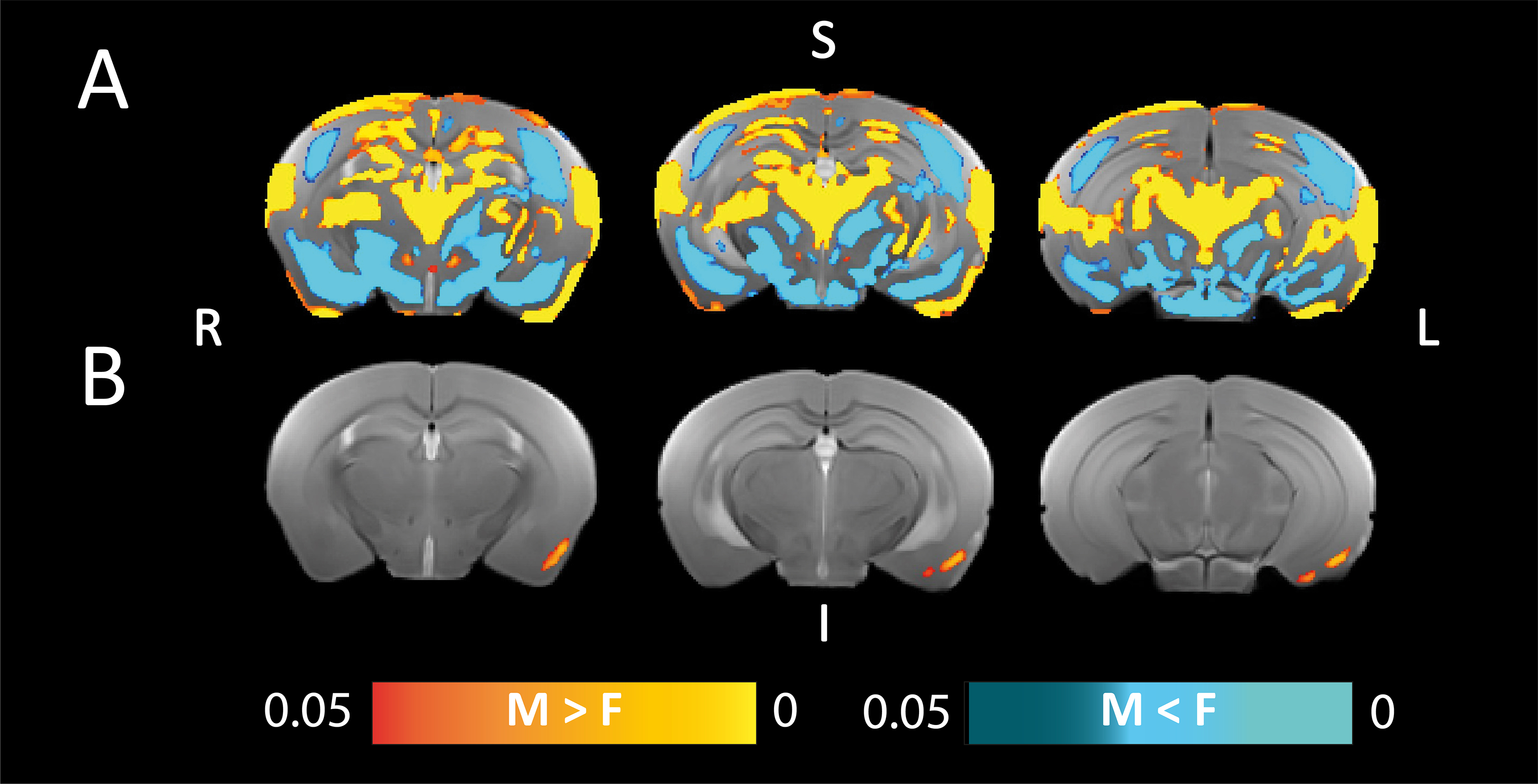
We have generated a mouse MRI 3D-brain template, with three contrasts: anatomical, i.e., T2W, and dw-based, i.e., FA and MD (Fig.1). The MouseX DW-ALLEN annotation, created with the AMBA CCFv3 annotation as a starting point, delineates the template with high accuracy and correspondence to the Allen Mouse Brain annotation (Fig.1D). We correlated ROI-specific MD values with each cell population present in the 3D Cell Atlas11. Correlation between cell count and MD signal was statistically significant only for microglia count (p-value=0.041, Fig.2) and not for the other cell types. We demonstrated the bias introduced by the use of a male-based 3D template. When comparing the voxel-wisedeformation across sexes to male-only template, widespread significant positive and negative differences were evidenced, which were not present when a template built with both sexes is used (Fig.3). Furthermore, we demonstrated on new mice longitudinal DW-MRI data that the use of male-only template detects spurious sex differences across 5 regions (4 grey matter and one white matter, Fig.4), which are not present when a template built with both sexes is used to register the data.DISCUSSION
With the generation of the MouseX DW-ALLEN Atlas including both sexes, we demonstrated that males and females undergo significantly different deformations when matching a single-sex template, and that using a single-sex template introduces spurious sex effects in the analysis. In addition, we proved the utility of registering MRI data to a consensus mouse brain atlas - the Allen Mouse Brain - already widely adapted for many neuroimaging fields10,14,15.CONCLUSIONS
The MouseX DW-Allen Atlas fills an important gap in preclinical imaging research, promotes the inclusion of both sexes and opens a promising scenario of collaborative and multi-approach science.Acknowledgements
No acknowledgement found.References
1. Ocañas SR, Pham KD, Cox JEJ, et al. Microglial senescence contributes to female-biased neuroinflammation in the aging mouse hippocampus: implications for Alzheimer’s disease. J Neuroinflammation. 2023;20:188.
2. Lynch MA. Exploring Sex-Related Differences in Microglia May Be a Game-Changer in Precision Medicine. Front Aging Neurosci. 2022;14. Accessed September 14, 2023.
3. Guneykaya D, Ivanov A, Hernandez DP, et al. Transcriptional and Translational Differences of Microglia from Male and Female Brains. Cell Rep. 2018;24(10):2773-2783.e6.
4. Garcia-Hernandez R, Cerdán Cerdá A, Trouve Carpena A, et al. Mapping microglia and astrocyte activation in vivo using diffusion MRI. Sci Adv. 2022;8(21):eabq2923.
5. Avants BB, Tustison NJ, Song G, Cook PA, Klein A, Gee JC. A reproducible evaluation of ANTs similarity metric performance in brain image registration. NeuroImage. 2011;54(3):2033-2044.
6. Jenkinson M, Beckmann CF, Behrens TEJ, Woolrich MW, Smith SM. FSL. NeuroImage. 2012;62(2):782-790.
7. Pasternak O, Sochen N, Gur Y, Intrator N, Assaf Y. Free water elimination and mapping from diffusion MRI. Magn Reson Med. 2009;62(3):717-730.
8. Fadnavis S, Batson J, Garyfallidis E. Patch2Self: Denoising Diffusion MRI with Self-Supervised Learning.
9. Leemans A, Jeurissen B, Sijbers J, Jones DK. ExploreDTI: A graphical toolbox for processing, analyzing, and visualizing diffusion MR data. 17th Annu Meet Intl Soc Mag Reson Med. 2009;17.
10. Wang Q, Ding SL, Li Y, et al. The Allen Mouse Brain Common Coordinate Framework: A 3D Reference Atlas. Cell. 2020;181(4):936-953.e20.
11. Erö C, Gewaltig MO, Keller D, Markram H. A Cell Atlas for the Mouse Brain. Front Neuroinformatics. 2018;12. Accessed September 18, 2023.
12. The control of the false discovery rate in multiple testing under dependency. Accessed October 23, 2023. https://projecteuclid.org/journals/annals-of-statistics/volume-29/issue-4/The-control-of-the-false-discovery-rate-in-multiple-testing/10.1214/aos/1013699998.full
13. Ashburner J, Hutton C, Frackowiak R, Johnsrude I, Price C, Friston K. Identifying global anatomical differences: deformation-based morphometry. Hum Brain Mapp. 1998;6(5-6):348-357.
14. Wang M, Zhuo L, Ma W, Wu Q, Zhuo Y, Wang X. AllenDigger, a Tool for Spatial Expression Data Visualization, Spatial Heterogeneity Delineation, and Single-Cell Registration Based on the Allen Brain Atlas. J Phys Chem A. 2023;127(12):2864-2872.
15. Moh ESX, Dalal S, Hutchinson MR, Packer NH. Mouse brain glycomics – Insights from exploring the Allen Brain Atlas and the implications for the neuroimmune brain. Brain Behav Immun. 2023;113:83-90.
Figures